Федеральное государственное бюджетное учреждение науки ИНСТИТУТ БИОЛОГИИ РАЗВИТИЯ им. Н.К. Кольцова РАН Koltzov Institute of Developmental Biology of Russian Academy of Sciences
Лаборатория ЭВОЛЮЦИОННОЙ ГЕНЕТИКИ РАЗВИТИЯ
Направления исследований
Основное направление исследований - изучение молекулярно-генетических механизмов формирования сложных морфологических и физиологических признаков, ранних этапов эволюции, механизмов формирования изолирующих барьеров между видами.
Исследования ведутся на всех уровнях организации живого – от молекулярно-генетического до морфологического и популяционного.
1. Концепция хромосомного видообразования: анализ роли перестроек и сопряженных с ними молекулярно-генетическим механизмов формирования изолирующих барьеров.
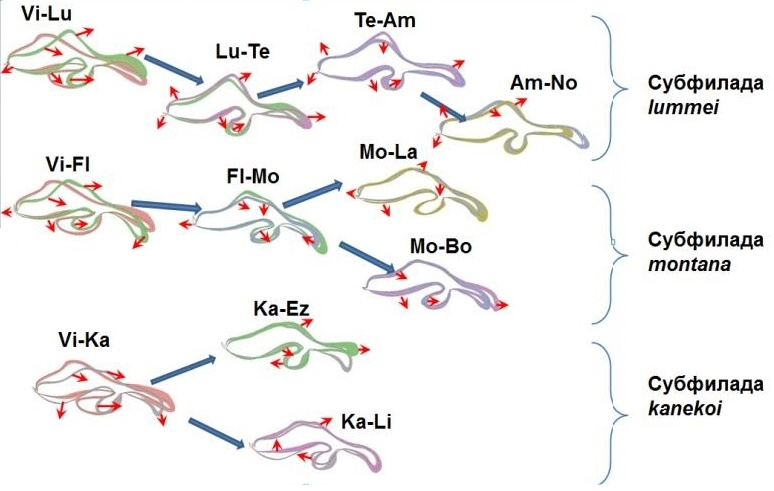
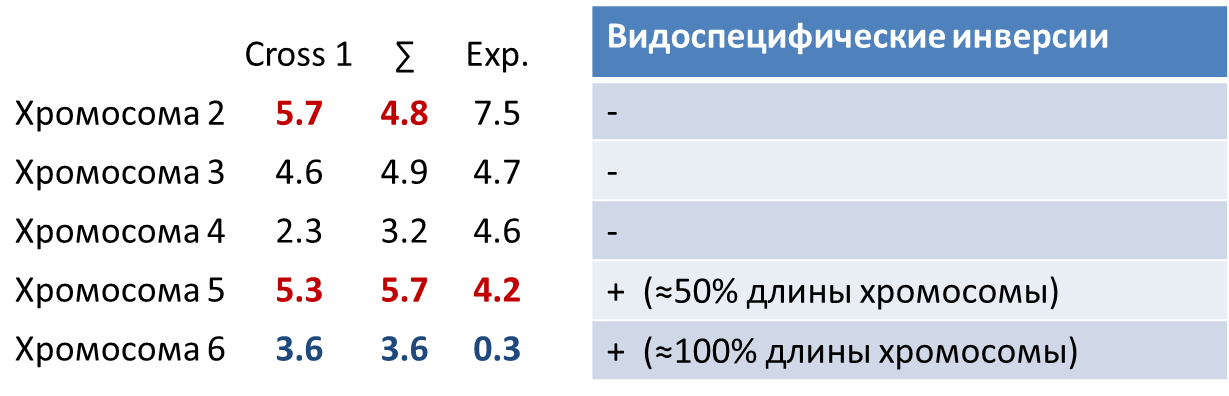
Анализ эволюционной роли перестроек у близкородственных видов дрозофил подтверждает справедливость хромосомной теории видообразования. В накоплении видоспецифической изменчивости видоспецифических признаков формы копулятивного аппарата (рис. 1.1) принимают участие все хромосомы, но основная роль принадлежит 5-й и 6-й аутосомам, несущим видоспецифические инверсии, что соответствует модели «подавления рекомбинации». (рис. 1.2)
2. Симбиоз как эволюционный фактор: от формирования эукариотической клетки до адаптивной иррадиации и дивергенции видов
Появлению эукариотической клетки должен был предшествовать более или менее длительный период коэволюции ее будущих компонентов в одном микробном сообществе, в течение которого между видами складывалась сложная система взаимоотношений и связей, необходимая для координации различных аспектов их жизнедеятельности. Молекулярные механизмы, развившиеся в ходе формирования этих синэкологических связей, могли сыграть важную роль в последующем процессе объединения нескольких прокариот в единую клетку. Анализу роли симбиотических отношений на ранних этапах эволюции и значению этого явления на современном этапе посвящен ряд исследований нашей лаборатории.
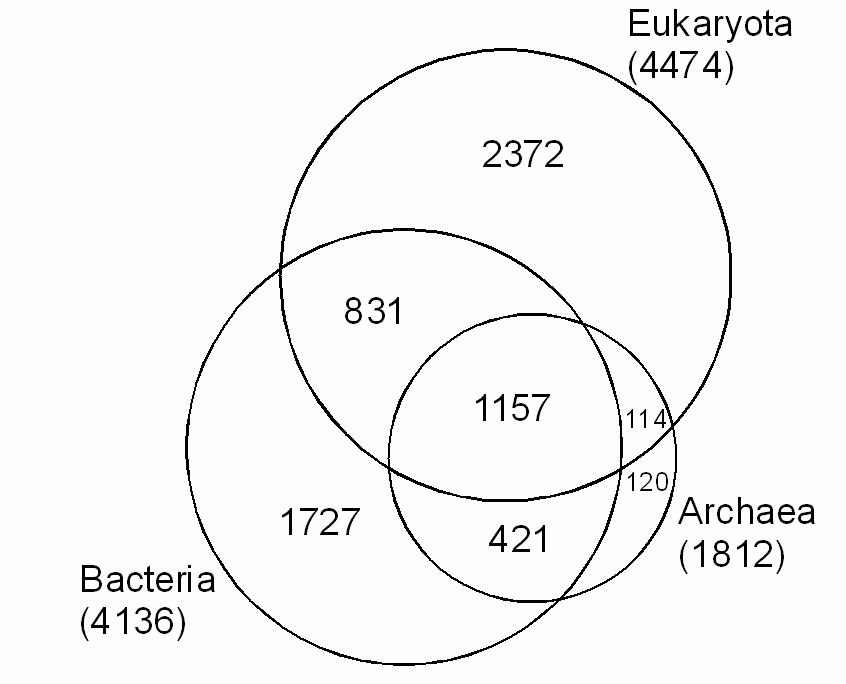
Анализ распределения белковых доменов (рис. 2), внесенных в 15-ую версию базы данных Pfam (август 2004 г.), в трех надцарствах: Archaea, Bacteria и Eykaryota, показал, что из общего числа белковых доменов эукариот почти половина была унаследована от прокариотических предков. От архей эукариоты унаследовали наиболее важные домены, связанные с информационными процессами нуклеоцитоплазмы (репликацией, транскрипцией, трансляцией). От бактерий унаследована значительная часть доменов, связанных с базовым метаболизмом и с сигнально-регуляторными системами. Очевидно, сигнально-регуляторные домены, общие для бактерий и эукариот, у первых выполняли синэкологические функции (обеспечение взаимодействия клетки с другими компонентами прокариотного сообщества), а у вторых стали использоваться для обеспечения согласованной работы клеточных органелл и отдельных клеток многоклеточного организма. Многие эукариотические домены бактериального происхождения (в том числе «синэкологические») не могли быть унаследованы от предков митохондрий и пластид, а были заимствованы у других бактерий. Предложена модель становления эукариотической клетки путем ряда последовательных симбиогенетических актов. Согласно этой модели, предком ядерно-цитоплазматического компонента эукариотической клетки была архея, у которой в условиях кризиса, вызванного ростом концентрации свободного кислорода в прокариотном сообществе, резко активизировался процесс инкорпорации чужеродного генетического материала из внешней среды (Марков, Куликов, 2005).
Роль симбиотических отношений на ранних стадиях дивергенции видов показана на модели формирования изолирующих барьеров между инфицированными и не инфицированными грамотрицательными альфа-протеобактериями Wolbachia линий Drosophila melanogaster (Куликов, Марков, 2007; Markov et al., 2009). Особенности эволюции бактериальных сообществ, вступающих в факультативные или облигатные симбиотические отношения с высшими организмами, проанализированы на примере цианобактерий (Koksharova et al., 2013; Кравцова и др., 2013) (рис. 2). Работа ведется в сотрудничестве с НИИ Физико-Химической Биологии им. А.Н. Белозерского МГУ.
Рис. 1.1. Эволюция признаков формы копулятивного аппарата дрозофил группы virilis.
Рис. 1.2.
Среднее значение объясненной изменчивости по признакам формы копулятивного аппарата на хромосому (с учетом размера групп скоррелированной изменчивости) из возвратных скрещиваний D.virilis x D.lummei Cross 1 – скрещивание ♂Xvir Ylum, Avir Alum x ♀Xvir Xvir, Avir Avir , доля объясненной изменчивости на хромосому;
∑ - суммарные данные от скрещиваний ♂Xvir Ylum, Avir Alum x ♀Xvir Xvir, Avir Avir и ♂Xlum Yvir, Alum Avir x ♀Xvir Xvir, Avir Avir, , доля объясненной изменчивости на хромосому;
Exp – ожидаемая доля изменчивости на хромосому
Рис. 2. Происхождение белковых доменов эукариотической клетки - гомология с доменами архей и бактерий
3. Механизмы и особенности молекулярной эволюции
Подтверждение неравномерности хода молекулярных часов в разных эволюционных родословных и для разных последовательностей ядерных генов получено для близнецовых видов дрозофил группы virilis с использованием тестов Таджимы и Такезаки (Куликов и др., 2010 а,б). С использованием авторской модели, объединяющей поправку Гилеспи и индекс инфляции Балмера, показано неравномерное накопление молекулярной изменчивости в пределах каждой эволюционной родословной дрозофил, обсуждаются причины, вызывающие неравномерный ход молекулярных часов (Куликов и др., 2010в). Точные оценки равномерности хода молекулярных часов по последовательностям митохондриальных и ядерных генов у видов дрозофил группы virilis свидетельствуют о нарушениях хода молекулярных часов в большинстве эволюционных линий данной группы видов. Замедление хода молекулярных часов характерно для поздних стадий дивергенции исследуемых видов.
В ходе эволюции молекулярных некодирующих последовательностей генома, в том числе области промотора и регуляторных последовательностей гена, могут происходить «мгновенные» эволюционные преобразования, за промежуток времени, соответствующий нескольким поколениям. Такие преобразования представляют собой полную замену значительных фрагментов последовательности, при быстром восстановлении функциональной активности гена. Анализ изменчивости регуляторной области гена Dras1 (рис. 3.1) и его ортологов у видов дрозофил разной степени родства из подродов drosophila и sophophora выявил неоднократную смену промотора, точки старта транскрипции и всей регуляторной области гена на разных ветвях филогенетического дерева дрозофил.
Изменения структуры последовательности связано с инсерциями мобильных элементов и в ряде случаев – с сопровождающими их структурными перестройками хромосомы. Исследуемая модель характеризуется строгим функциональным и структурным консерватизмом кодируемого геном белка, и летальным характером 0-аллелей, исключающим экспрессию данного гена. Транспозиции мобильных элементов в область промотора приводят к формированию аллелей гена с летальным эффектом или, по крайней мере, с резко сниженными показателями жизнеспособности. В природе такие аллели сохраняются исключительно в гетерозиготном состоянии и теряются вследствие генетико-автоматических процессов. Отмеченная смена промотора и регуляторной части должна сопровождаться восстановлением функциональной активности гена в течение ограниченного числа поколений.
Комплексные исследования молекулярных и морфологических признаков родственных видов дрозофил показали, что преобразования кариотипа, белков, нуклеотидных последовательностей ДНК и морфологических признаков могут происходить независимо и с неодинаковой скоростью, что приводит к разным эволюционным последствиям. Так, сопоставляя закономерности эволюции количественных признаков формы копулятивного аппарата и молекулярной изменчивости можно заключить, что неравномерная скорость накопления эволюционно-значимой и нейтральной изменчивости опосредована действием различных эволюционных факторов. Нарушения скорости накопления изменчивости по статусным видовым признакам связаны с действием полового отбора, выступающего в роли дизруптивного в момент расхождения видов. Нарушения скорости накопления нейтральной изменчивости вызваны действием генетико-автоматических процессов.
В работах по анализу эволюционно значимой изменчивости признаков копулятивного аппарата дрозофил показано, что степень доминирования нового фенотипа усиливается с течением времени, определяя темп и направленность накопления изменчивости; эволюция доминантности осуществляется за счет эпистатической изменчивости, в соответствии с концепцией Фишера «эволюции доминантности».
Применен новый методологический подход, позволяющий проводить исследование генетического разнообразия предковых популяций в условиях ограниченных выборок на материале одного образца геномного сиквенса. Анализ геномных данных показал, что в ядерном геноме D. virilis содержатся многократные копии фрагментов мт-геномов (а также целых мт-геномов), так называемые Numt-последовательности. Было выявлено 16 копий фрагмента гена atp6 длиной 520 п.о. мт-генома D. virilis. Выявлено 7 независимых событий переноса мт-фрагментов в ядерный геном и реконструированы мт-гаплотипы, давшие начало митохондриальным псевдогенам. C помощью парсимониальной дендрограммы определен спектр мт-гаплотипов предковой популяции D. virilis (рис. 3.2), их возраст и возраст событий переноса фрагментов мтДНК в ядерный геном. Самое раннее событие переноса соответствует 720 тыс. л. назад. Дивергенция мт-гаплотипов началась около 1 млн. л. назад. Средняя величина значений попарных генетических расстояний архаичных мт-гаплотипов (p) составляет 0,044. Данные согласуются с данными анализа генетического разнообразия природных популяций других видов дрозофил группы virilis, в отличие от разнообразия современных синантропных популяций D.virilis, генетический полиморфизм которых значительно обеднен.
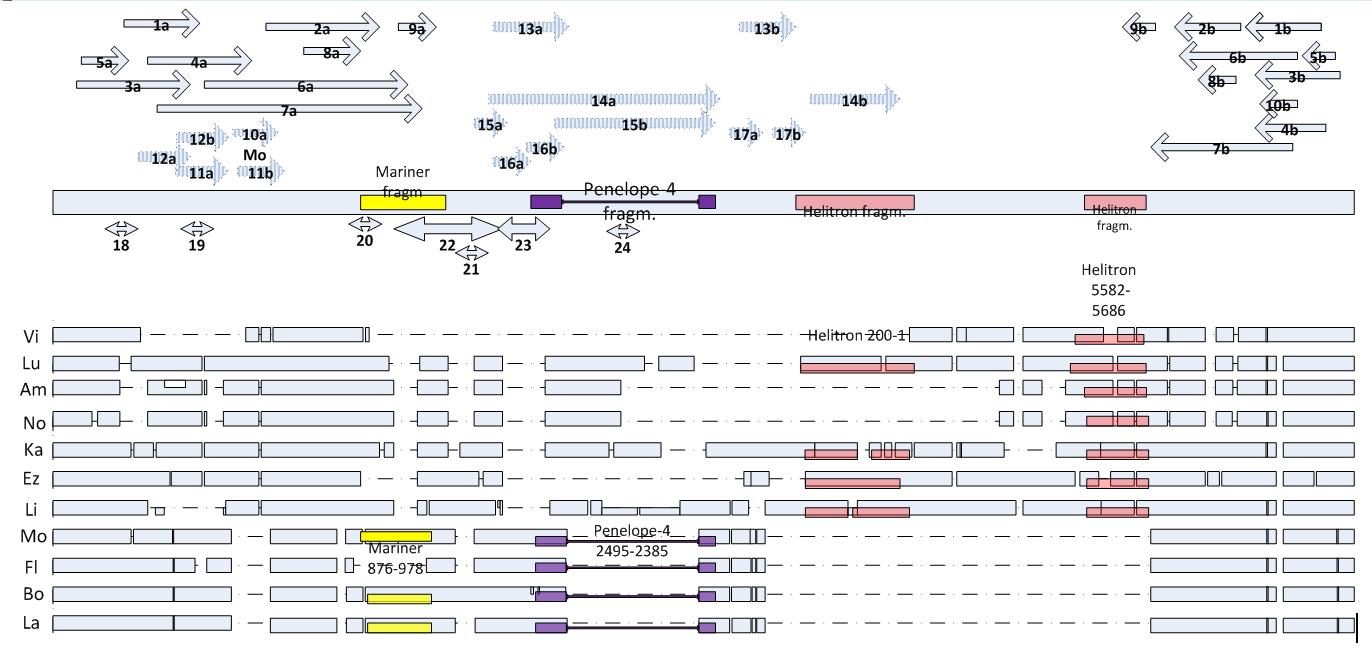
Рис. 3.1. Детализация структуры области промотора области межгенного спейсера, расположенного выше Dras1 гена у видов группы virilis. а. Положение повторов и палиндромов; б. распределение крупных делеций и фрагментов МЭ.
а) Гомологичные повторы имеют идентичные номера с буквенным обозначением a, b. Наличие повтора у данного вида указано двухбуквенной аббревиатурой. Inverted repeats (сплошные стрелки): 1a,b – Mo, La, Lu, Ez, Li; 2a,b – Li, Vi, Am; 3a.b – Ka; 4a,b – Am; 5a,b – Vi; 6a,b – Ez, Lu; 7a,b – Bo; 8a,b – La, Mo; 9a,b – La, Am; 10a,b – Mo. Direct repeats (прерывистые стрелки): 11a,b – Ez, Mo, La, Bo; 12a,b – Lu; 13a,b – La, Mo; 14a,b – Ka; 15a,b – Bo; 16a,b – Bo; 17a,b – Li. Palindromes (двунаправленные стрелки): 18 – Ez; 19 – Bo, Ka; 20 – La, Mo, Bo, Ez; 21 – Am, Ka; 22 – Li; 23 – Bo; 24 – Ka.
б) прямоугольные серые блоки – последовательность ДНК, прерывистые линии и разрывы между блоками – гэпы, двукратное уменьшение ширины серых блоков – отмеченный внутривидовой полиморфизм по гэпам. Цветными блоками отмечены фрагменты, проявляющие гомологию с фланкирующими последовательностями мобильных элементов.
Рис. 3.2. Numts и оценка биоразнообразия предковых популяций
4. Молекулярная систематика и филогения, гибридизация в природе
Изучена изменчивость последовательности гена kl-2 1-beta dynein Heavy chain у 11 видов дрозофил группы virilis. Благодаря отсутствию рекомбинации генов, локализованных на Y-хромосоме анализ позволяет провести сравнение материнской и отцовской генеалогий. Минимальное число нуклеотидных замен на сайт (0,005) зафиксировано между видами D.a.americana и D.a.texana. Среднее значение попарных генетических дистанций в группе составляет 0,036. Различий между последовательностями kl-2 у разных лабораторных линий D.virilis не было выявлено. Напротив, последовательности kl-2 двух линий D.littoralis из южнокавказской популяции и из североевропейской популяции отличаются. Южная популяция является предковой по отношению к северной. Скорость фиксации нуклеотидных замен сильно отличается у разных видов, возможно, из-за различий в численности популяций. Маленькие островные популяции D.kanekoi из Японии эволюционируют значительно быстрее, чем большие континентальные популяции D.littoralis. В отличие от данных мтДНК, отражающих генеалогию по материнской линии, данные У-хромосомы указывают на то, что D.kanekoi и D.ezoana образуют один кластер. Таким образом, можно предполагать, что в истории этих видов были этапы несимметричной гибридизации, приведшие к различиям в генеалогиях по материнской и отцовской линиям.
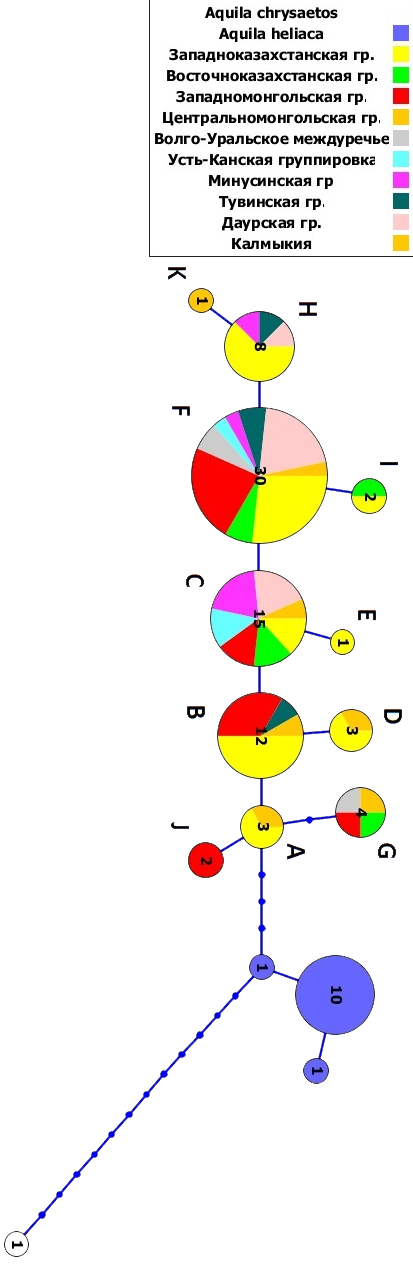
Ведется работа по оценке генетического разнообразия популяций степного орла Aquila nipalensis в условиях быстрого сокращения численности вида (рис. 4). Впервые получены нуклеотидные последовательности D-петли мтДНК степного орла. Общая длина фрагмента составляет 1162 п.о. Проанализировано более 100 особей из различных участков ареала обитания данного вида. Показан полиморфизм по фрагменту D-петли мтДНК, выявлено 11 мт-гаплотипов, 4 из которых – мажорные. Для вида характерно существование в виде малочисленных популяций с периодическими колебаниями численности, приводящими к сменам частот гаплотипов. В малочисленных популяциях данного вида отмечены случаи гибридизации с орлом-могильником. Степной орел, вид с низким уровнем генетического разнообразия, даже без участия человеческого фактора находится под угрозой внезапного исчезновения. Это угрожаемый вид, мониторинг состояния которого должен вестись постоянно.
Рис. 4. Степной орел – вид с низким генетическим разнообразием.
Наиболее древний гаплотип A и его ближайшие производные B и J встречаются только в отдельных популяциях: Калмыцкой и Западноказахстанской на западе, и Западномонгольской и Тувинской на востоке – предположительных центрах видообразования. Не выявлено специфических групп гаплотипов для западных и восточных популяций.
5. Генетические основы количественных признаков, участвующих в формировании изолирующих барьеров между видами.
Анализ закономерностей формирования изменчивости (рис. 5.1) количественных морфологических признаков проведен на признаках формы копулятивного аппарата самцов и формы крыловой пластины дрозофил группы virilis. Было показано, что на протяжении всего времени дивергенции видов группы virilis форма копулятивного органа самцов находилась под действием направленного полового отбора. Максимальному давлению отбора подвержены признаки дорсальной поверхности эдеагуса, шипа на его конце и формы парамеров (Куликов и др., 2008). Скорость накопления видоспецифической изменчивости по признакам формы копулятивного аппарата различается как между разными эволюционными линиями дрозофил группы virilis, так и в пределах каждой родословной, и связь эволюционно-значимых частей копулятивного аппарата дрозофилы с половым поведением опосредована сенсорными микрохетами (рис. 5.1) на дорсальной поверхности эдеагуса (Куликов и др., 2013). Эволюция статусных признаков формы копулятивного аппарата у дрозофил осуществляется на основе сохраняющейся генетической изменчивости, в соответствии с концепцией Орра «Сито Холдейна и адаптации на основе поддерживающейся в популяции
генетической изменчивости».
Эволюция формы крыла у близнецовых видов virilis не имеет направленного характера, связана преимущественно с аллометрическими эффектами и указывает на участие данного признака в формировании частных адаптаций (рис. 5.2). Дивергенция по форме крыловой пластины осуществляется значительно медленнее, чем по признакам
формы полового аппарата.
Numt-последовательности, или ядерные последовательности митохондриального происхождения, позволяют оценить время события, в результате которого формируется такая последовательность, и используются для маркировки эволюционных родословных общего и независимого происхождения. На модели клеточных культур дрозофилы изучены механизмы возникновения numt-последовательностей (Андрианов и др., 2013).
6. Пре– и постзиготические механизмы формирования изоляционных барьеров при видообразовании
Обсуждая механизмы изоляции, принято выделять презиготические, в том числе пред- и посткопуляционные, и постзиготические.
С использованием видов-двойников дрозофил группы virilis (рис. 6.1) изучается роль и генетические механизмы формирования предкопуляционных изолирующих барьеров, связанных с признаками копулятивного аппарата самцов дрозофил (Куликов и др., 2013 а,б) и особенностями брачного поведения (Vedenina et al., 2013). Форма копулятивного аппарата самцов близкородственных видов имеет качественные отличия, внутривидовая изменчивость, как правило, существенно ниже межвидовой, и локусы, определяющие видоспецифические различия, локализованы во всех хромосомах. Анализ брачного поведения дрозофил группы virilis (рис. 6.2) выявил характерные особенности поведенческих программ брачного поведения, заключающиеся в продолжительности и очередности отдельных элементов ритуала.
Используя разные способы блокирования обонятельных, осязательных и вкусовых рецепторов на антеннах, конечностях и крыльях дрозофилы (рис. 6.3), удалось установить зависимость между рецепцией сигнала от партнера и последовательностью отдельных элементов ухаживания. В гетероспецифических скрещиваниях наблюдается «сбой» поведенческих программ, связанный не только с изменением звуковых стимулов (характер частотно-амплитудных колебаний крыльев обоих партнеров), но и с изменением обонятельных и вкусовых стимулов. Такие нарушения приводят к затягиванию отдельных элементов ритуала, задержке или неспособности перейти к следующей стадии ухаживания, и в конечном итоге к многократному удлинению времени ухаживания, сокращению периода копуляции (и качества осеменения, соответственно), или к полной неспособности к копуляции.
Работы по анализу постзиготических механизмов формирования изолирующих барьеров описаны в разделе «История лаборатории», в части истории лаборатории Генетики.
7. Молекулярно-генетические механизмы формирования сложных морфологических и физиологических признаков
(механизмы детерминации пола, признаки направленной асимметрии, формирование пищевых адаптаций, хозяйственно-ценные признаки, генетические основы агрессивного
поведения)
В рамках изучения генетических основ количественных признаков, связанных с хозяйстенно-ценными характеристиками сельскохозяйственных и промысловых животных, в сотрудничестве с ИОГЕН РАН (лаб. сравнительной генетики животных) проводится работа по выявлению молекулярно-генетических маркеров эффективности селекции по признакам качества молочной и мясной продукции крупного рогатого скота (Столповский и др., 2010; Лазебная и др., 2010; Лазебная и др., 2011), качества меха соболя (Каштанов и др., 2011а; Нюхалов и др., 2012), а так же геногеографии природных и доместицированных популяций соболя (Каштанов и др., 2010; Каштанов и др., 2011б; Андрианов и др., 2012; Рожнов и др., 2013).
Совместно с кафедрой Теории эволюции МГУ ведутся работы по анализу генетических механизмов формирования пищевых адаптаций модельного вида Drosophila melanogaster к условиям повышенной солености, изменению состава корма по углеводам (замена сахара на крахмал), полулетальным дозам инсектицидов.
Особый интерес представляет вопрос об эволюционном формировании признаков направленной асимметрии. Г.П. Удалова в обзоре, посвященном асимметрии беспозвоночных животных (Удалова, 2009) показала, что структурная асимметрия часто связана с функциональной асимметрией. Следовательно, можно предположить, что причины нарушения симметрии имеют адаптивную значимость и подвержены влиянию отбора. Однако анализ асимметричных билатеральных признаков животных показывает, что далеко не во всех случаях существование устойчивой направленной асимметрии можно объяснить непосредственным действием отбора. Традиционно, для экспериментального анализа механизмов в подобных случаях обращаются к животным моделям. Крылья дрозофил разных видов имеют направленную морфологическую асимметрию. Ее удалось показать с использованием методов геометрической морфометрии Букштейна, Рольфа и Маркуса. Как по нашим данным, полученным для 11 близнецовых видов дрозофил группы virilis, и в экспериментах с линиями D.melanogaster, так и по немногочисленным пока литературным источникам (Klingenberg et al., 1998; обзор Pelabon, Hansen, 2008) подтверждается наличие слабой, но достоверной направленной асимметрии крыльев у большинства исследованных видов насекомых. Формирование этой асимметрии тоже необъяснимо с позиции отбора (Pelabon, Hansen, 2008). Выявляемые области крыловой пластины, проявляющие признаки направленной асимметрии, достаточно сходны у разных видов и в разных условиях влияния внешних факторов. При этом степень изменчивости, опосредованной направленной асимметрией, весьма мала, и направление вектора изменчивости неоднозначно. Вместе с тем известно, что направленная асимметрия крыла – эволюционно-консервативный признак, не подверженный действию отбора (Ashley et al., 2009). Таким образом, предположение о влиянии внешних факторов на формирование признака направленной асимметрии формы крыла к дрозофилы пока еще является слабо обоснованным.
Рис. 5.1. Сенсорные микрохеты, определяющие действие полового отбора на форму копулятивного аппарата дрозофил.
Показано эволюционное происхождение микрохет копулятивного аппарата от микрохет абдомена насекомых. На снимке отдельной микрохеты отчетливо видна внутренняя полость, характерная для сенсорных микрохет. На эдеагусе микрохеты распределены на наиболее эволюционно изменчивых частях органа.
Рис. 5.2. Адаптивная изменчивость компартментов крыла дрозофил группы virilis.
У дрозофил с большими крыльями наблюдается уменьшение проксимальной области крыла и увеличение дистальной, наиболее активной в полете.
а. Зоны независимой изменчивости крыловой пластины дрозофил.
b. Аллометрические деформации формы крыловой пластины, полученные при оценке межвидовой изменчивости.
Рис. 6.1. Drosophila virilis
Рис. 6.2. Копулирующая пара Drosophila virilis
Рис. 6.3. Варианты микрохирургического удаления тарзусов и крыльев у самцов дрозофил
8. Генетические основы признаков агрессивности у человека
Комплексный междисциплинарный проект по изучению генетических основ девиантного агрессивного поведения у человека проводится в сотрудничестве с Институтом этнологии и антропологии имени Н. Н. Миклухо-Маклая РАН (Сектором кросс-культурной психологии и этологии человека) и Институтом биологии гена (Лабораторией организации генома). Анализ ассоциаций аллельного полиморфизма генов – кандидатов с выраженностью признаков агрессивного поведения и с антропометрическими признаками, связанными с маскулинным поведением, проводится на материале, собранном в обществах с традиционной племенной культурой, имеющими разный уровень культурно допустимой агрессии, в частности хадза и датога, (рис. 8.1-2) проживающих на севере Танзании. Выбор представителей доидустриальных обществ обусловлен стабильным состоянием культурных отношений, под действием которых на протяжении тысячелетий формировались социальные рамки допустимого агрессивного поведения. Показано участие дофаминовой, серотониновой и андрогеновой сигнальных систем в формировании признаков агрессивного поведения, и различный вклад генов, участвующих в передаче сигнала в данных системах, у представителей культур с разным уровнем допустимой агрессии (Butovskaya et al., 2012; Butovskaya M. et al., 2013; Васильев и др., 2011; Шибалев и др., 2013; Суходольская и др., 2014, Васильев и др., 2014). Аналогичный подход оценки ассоциации аллельного состояния генов, признаков агрессивности и формальных показателей профессиональной успешности применен при оценке связи уровня агрессии с профессиональными достижениями спортсменов (Butovskaya P. et al., 2013). В последних исследованиях показана положительная корреляция между активностью андрогеновой сигнальной системы, уровнем агрессивности и репродуктивным успехом у мужчин хадза и датога.
Рис. 8.1. Датога - праздник
Рис. 8.2. Датога - праздничные наряды
9. Биоинформатика: межмолекулярные взаимодействия в клетках и тканях, анализ данных, полученных высокопроизводительными методами
Формирование любого фенотипического признака, не говоря уже о фенотипе в целом – результат реализации информации, записанной в сотнях генов. Реализация осуществляется через контроль процессов роста, деления, дифференцировки и перемещения клеток и тканей, взаимодействия сотен и тысяч элементарных процессов, связанных в сложнейшую динамическую сеть межмолекулярных взаимодействий. Поэтому принципиальная задача биологии вообще, и генетики в частности – расшифровка пути от гена к признаку, не может быть решена без учета межгенных взаимодействий. В рамках инициативного проекта по разработке базы данных межмолекулярных взаимодействий, моделирования генных сетей и создания он-лайн доступного инструмента анализа и визуализации данных пользователей (геномные и экспрессионные данные, полученные с применением NGS-методов) разрабатывается и уже применяется
аналитическая платформа, включающая модули построения генных сетей (рис. 9), внесения данных пользователей, визуализации данных и алгоритмов обработки полученных результатов (выявления подсетей и отдельных регуляторных каскадов, оценки по данным онтологии генов клеточных, тканевых, онтогенетических, метаболических, патологических процессов и др.) (Shashova et al., 2014; Сорокина и др., 2013). Ряд исследований, в которых принимали участие сотрудники, посвящен экспериментальной, статистической и биоинформационной оценке механизмов внутриклеточных регуляций и формирования патологий (Ekins et al., 2006 a,b; Зайкова и др., 2011).
В рамках научно-исследовательской работы по теме лаборатории совместно с ООО «СибЭкоЦентр» сотрудники ИБР РАН принимают участие в экспедициях по мониторингу, кольцеванию и мечению хищных птиц (рис. 10.1), а также ведут работы по поддержке и расширению
уникальной коллекции линных перьев хищных
птиц для молекулярно-генетического анализа (рис. 10.2. a-b).
В ходе исследований механизмов формирования изоляционных барьеров при видообразовании у птиц сотрудники лаборатории эволюционной генетики развития ИБР РАН приняли участие в пилотном проекте по реинтродукции уникальной морфы сокола балобана путем подсаживания искусственно выращенных птенцов в естественные гнезда других морф пернатого хищника. (видео 10.1.)
Видео 10.1. Репортаж телеканала Россия 1 "Вести" от 27.06.2017