 © Кремнёв Станислав Валерьевич,
кандидат биологических наук, старший научный сотрудник, лаборатория эволюции морфогенезов ИБР РАН. Научные интересы: морфогенез, эволюция эмбрионального развития.
© Кремнёв Станислав Валерьевич,
кандидат биологических наук, старший научный сотрудник, лаборатория эволюции морфогенезов ИБР РАН. Научные интересы: морфогенез, эволюция эмбрионального развития. В любом учебнике по биологии развития можно найти информацию о том, что для формирования нервной трубки эмбриону любого позвоночного необходим индукционный сигнал от зачатка хорды, обладающего свойствами региона организатора. Более того, существует "классическая" модель, в которой сигнал зачатка хорды имеет решающее значение не только для спецификации дна нервной трубки, но и для ее дорзо-вентральной разметки (Ribes and Briscoe, 2009). Роль осевой мезодермы в дорзо-вентральной разметке нервной трубки связана с секрецией сигнальной молекулы Sonic hedgehog (Shh), запускающей экспрессию того же гена в клетках дна нервной трубки (рис. 1, А).
Однако универсальность функции хорды как организатора остается под вопросом. У рыбки Данио (Danio rerio) ни удаление хорды микрохирургическим путем, ни нарушение формирования хорды у мутантов no tail не приводят к нарушениям дифференцировки дна нервной трубки. У шпорцевой лягушки (Xenopus laevis), активность Shh слабо регулирует экспрессию генов в дне нервной трубки (Peyrot et al., 2011). Также имеются противоречивые данные об индукторе спецификации дна нервной трубки у птиц (Teillet et al., 1998).
Несмотря наличие таких данных, до сих пор не была подробно исследована динамика экспрессии shh, начиная с самых ранних стадий развития, у представителей разных групп позвоночных животных. Работа, опубликованная в журнале EvoDevo в 2018 году С.В. Кремнёвым и соавторами, как раз и была направлена на заполнение данного пробела (Kremnyov et al., 2018).
Её результаты поставили под сомнение универсальность функции хорды как региона-организатора позвоночных.
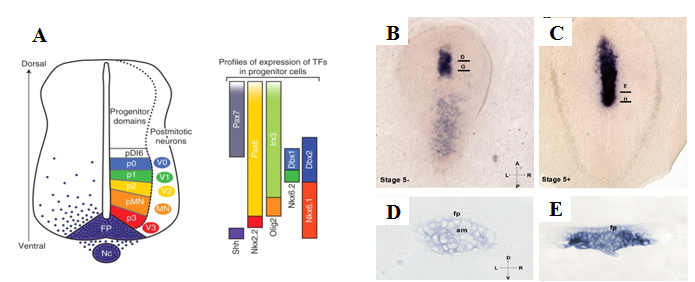
Рис.1 A. Классическая схема индукции shh в клетках дна нервной трубки и ее дорсовентральной разметки (Ribes and Briscoe, 2009). B-E. Пространственная динамика экспрессии shh во время гаструляции кролика. A. Стадия 5-, В. Стадия 5+. D-E. Поперечные срезы. Динамика экспрессии соответствует представлениям классической модели. fp – дно нервной трубки, am – осевая мезодерма
Оказалось, что у кролика экспрессия Shh впервые детектируется в зачатке хорды, и только потом обнаруживается в области дна нервной трубки, что полностью согласуется с общепринятой моделью индукции этой структуры (рис. 1, B-E).
Изучение паттерна экспрессии Shh у эмбрионов птиц показало, что на стадиях формирования и морфогенеза хорды Shh экспрессируется полосой вдоль серединной линии зародыша (рис. 2, A-C). При исследовании паттерна экспрессии на срезах было показано, что Shh в нервной пластинке детектируется на высоком уровне в задних отделах формирующейся нервный системы, непосредственно перед гензеновским узелком (рис. 2, D, E). А именно в этом регионе экспрессия Shh в подлежащей хорде отсутствует. Такой паттерн экспрессии Shh у эмбриона птицы ставит под сомнение представления классической модели об индукционном механизме предразметки нервной трубки.
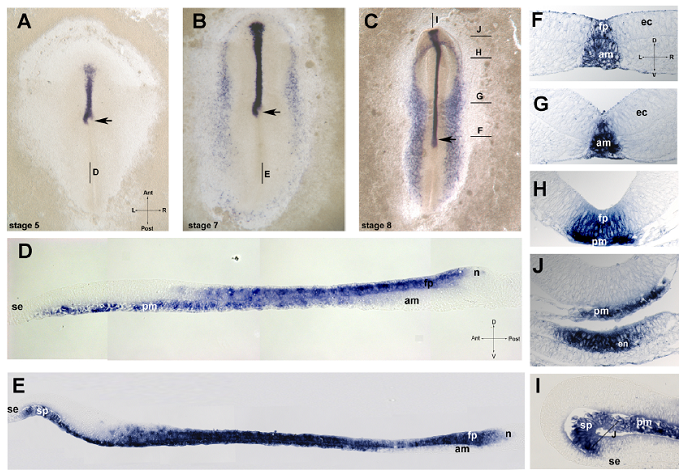
Рис. 2. Пространственная динамика экспрессии shh в развитии курицы. A-C: тотальные препараты эмбрионов на стадии 5 (А), 7 (B) и 8(С); сагиттальный срез эмбриона на стадии 5 (D) и стадии 7 (E); F-J поперечные срезы эмбрионов на стадии 8 HH на уровнях указанных на C. I – сагиттальный срез эмбриона на стадии 8. Обозначения: fp – дно нервной трубки, am – осевая мезодерма, ec – эктодерма, en – эндодерма, n – узелок, pm – прехордальная мезодерма, sp – преоральная область кишки, se – поверхностная эктодерма, стрелка – положение гензеновского узелка, Пересекающиеся стрелки указывают анатомические оси: A – anterior, P – posterior, L – лево, R – право, D – дорзальный, V – вентральный.
Сразу встаёт вопрос, когда начинается экспрессия shh в зачатках хорды и дна нервной трубки? Оказалось, что сигнал Shh в области будущего дна нервной трубки детектируется очень рано – ещё до формирования хорды и даже до начала гаструляции (рис. 3). Таким образом, классическая модель дорсовентрвльной разметки нервной трубки не может быть применена к развитию птиц.
Авторы также исключили возможность того, что функцию разметки нервной трубки у птиц на себя взял другой представитель семейства hedgehog: ihh или dhh. Экспрессия ни Ihh, ни Dhh, не была детектирована ни в хорде, ни в дне нервной трубки на исследованных стадиях развития эмбриона птицы.
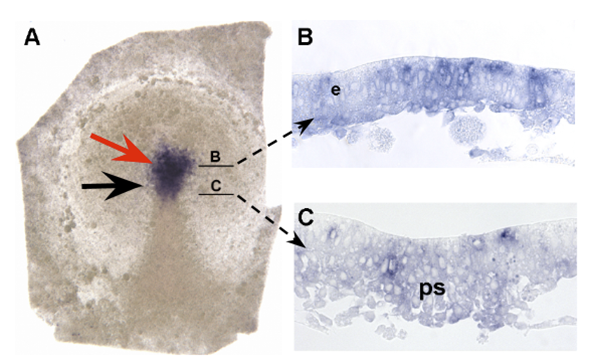
Рис. 3. Ранняя экспрессия shh в эмбрионе курицы на стадиях 2+/3- HH. Чёрной стрелкой указана антериорная граница первичной полоски, красной – экспрессия shh в области будущего дна нервной трубки.
Можно заключить, что авторы статьи выявили значительные различия в динамике экспрессии shh у представителей разных классов позвоночных (рис. 4). Это указывает на эволюционную дивергенцию механизмов индукции дна нервной трубки. Вероятно, именно эволюционная пластичность ранней эмбриональной индукции позволила позвоночным в широких пределах изменять гаструляционные морфогенезы в связи с эволюцией репродуктивных стратегий, которая затрагивала организацию яйцеклетки.
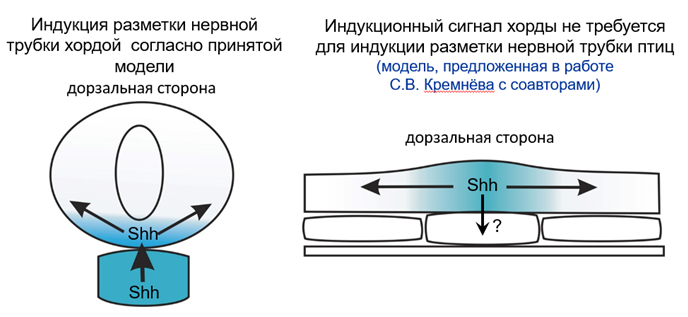
Рис. 4. Схема поперечного среза эмбриона курицы на уровне постериорной хорды. A: вентродорзальная индукция дифференцировки дна нервной трубки согласно принятой модели – shh изначально начинает экспрессироваться в хорде, а далее индуцирует экспрессию shh в дне нервной трубки. Белок Shh из обоих центров формирует градиет. B: изменённая модель демонстрирующая раннюю экспрессию shh в дне нервной трубки и формирование градиента shh в пределах нервной пластинки. Стрелка с вопросительным знаком предполагает индукцию shh в хорде клетками дна нервной трубки.
Результаты данной работы выявляют множество нерешенных вопросов и определяют направление дальнейших исследований. Важным вопросом является выяснение молекулярных путей, запускающих и регулирующих изначальную экспрессию shh в хорде у млекопитающих и дне нервной трубки у птиц. Являются ли эти пути консервативными у разных групп животных и если да, то что привело к изменению локализации их активности? Интересным вопросом является и то, когда именно произошла дивергенция механизмов разметки нервной трубки позвоночных, а также когда возникли индукционные взаимодействия между хордой и формирующейся нервной системой. Для ответа на эти вопросы необходимо включить в исследование представителей других классов позвоночных животных, например, рептилий. Исключительно важным вопросом является и выяснение того, когда именно у разных групп животных происходит спецификация тех областей нервной трубки, из которых в последствии дифференцируются нейроны разных типов и нужен ли вообще для их дифференцировки установление градиента shh.
Kremnyov, S., Henningfeld, K., Viebahn, C. and Tsikolia, N., 2018. Divergent axial morphogenesis and early shh expression in vertebrate prospective floor plate. EvoDevo, 9(1), p.4.
Ribes V, Briscoe J. 2009, Establishing and interpreting graded Sonic Hedgehog signaling during vertebrate neural tube patterning: the role of negative feedback. Cold Spring Harb Perspect Biol., 1(2):a002014.
Peyrot, S.M., Wallingford, J.B. and Harland, R.M., 2011. A revised model of Xenopus dorsal midline development: differential and separable requirements for Notch and Shh signaling. Developmental biology, 352(2), pp.254-266.
Teillet, M.A., Lapointe, F. and Le Douarin, N.M., 1998. The relationships between notochord and floor plate in vertebrate development revisited. Proceedings of the National Academy of Sciences, 95(20), pp.11733-11738.
Новость подготовил © КРЕМНЁВ С.В.
29.04.2019