Martynov A., Lundin K., Picton B., Fletcher K., Malmberg K., Korshunova T. Multiple paedomorphic lineages of soft-substrate burrowing invertebrates: parallels in the origin of Xenocratena and Xenoturbella//PloS One. – 2020. – Vol. 15. – Is. 1. – P. e0227173. Diversity 2024, 16, 220. DOI: 10.1371/journal.pone.0227173.
Schiffer P.H., Natsidis P., Leite D.J., Robertson H.E., Lapraz F., Marlétaz F., Fromm B., Baudry L., Simpson F., Høye E., Zakrzewski A.C., Kapli P., Hoff K.J., Müller S., Marbouty M., Marlow H., Copley R.R., Koszul R., Sarkies P., Telford M.J. Insights into early animal evolution from the genome of the xenacoelomorph worm Xenoturbella bocki//Elife. – 2024. – Aug 7; 13:e94948. doi: 10.7554/eLife.94948

Область научных интересов: нейроэтология, нейробиология, нейротрансмиттеры, поведение.
Педоморфоз (утрата потомками взрослой стадии в процессе эволюции с приобретением черт более ранних стадий индивидуального развития предков) можно охарактеризовать, как процесс, связывающий онтогенез, эволюцию, и систематику. Одним из факторов, способствующих педоморфной эволюции морских беспозвоночных животных, является среда мягких субстратов, поскольку маленький размер (характерный для ряда педоморфных животных) и редукция выступающих частей тела, облегчают передвижение внутри частиц песка и рытье нор, и таким образом способствуют формированию новых трофических и нишевых адаптаций, появлению новых способов защиты от хищников. Предположение, что мягкие субстраты морских осадочных пород способствуют педоморфной эволюции морских беспозвоночных, никогда строго не проверялось с использованием молекулярно-филогенетических данных с широкой выборкой таксонов.
В работе Martynov et al., 2020 на основе 5 генов-маркеров проведено молекулярно-филогенетическое исследование представителей 21 семейства одной из крупных групп голожаберных моллюсков – Aeolidacea, некоторые представители которых встречаются на мягких субстратах (рис. 1). Представители одного из семейств Pseudovermidae (псевдочерви), обитающие среди частичек грунта, внешне похожи на червей, хотя у них сохранились рудиментарные спинные папиллы (рис. 2). Внутреннее строение Pseudovermidae, благодаря наличию радулы, челюстей, и ряда других признаков, однозначно указывает на их принадлежность к голожаберным моллюскам. Анализ показал, что представители семейства Pseudovermidae с четко выраженными педоморфными чертами являются сестринской группой (рис. 1) семейства Cumanotidae (которые еще имеют сложные морфологические признаки, но уже способны факультативно зарываться в мягкие субстраты). Также отмечено, что семейства Cumanotidae и Pseudovermidae сопоставимы по общим признакам строения радулы.
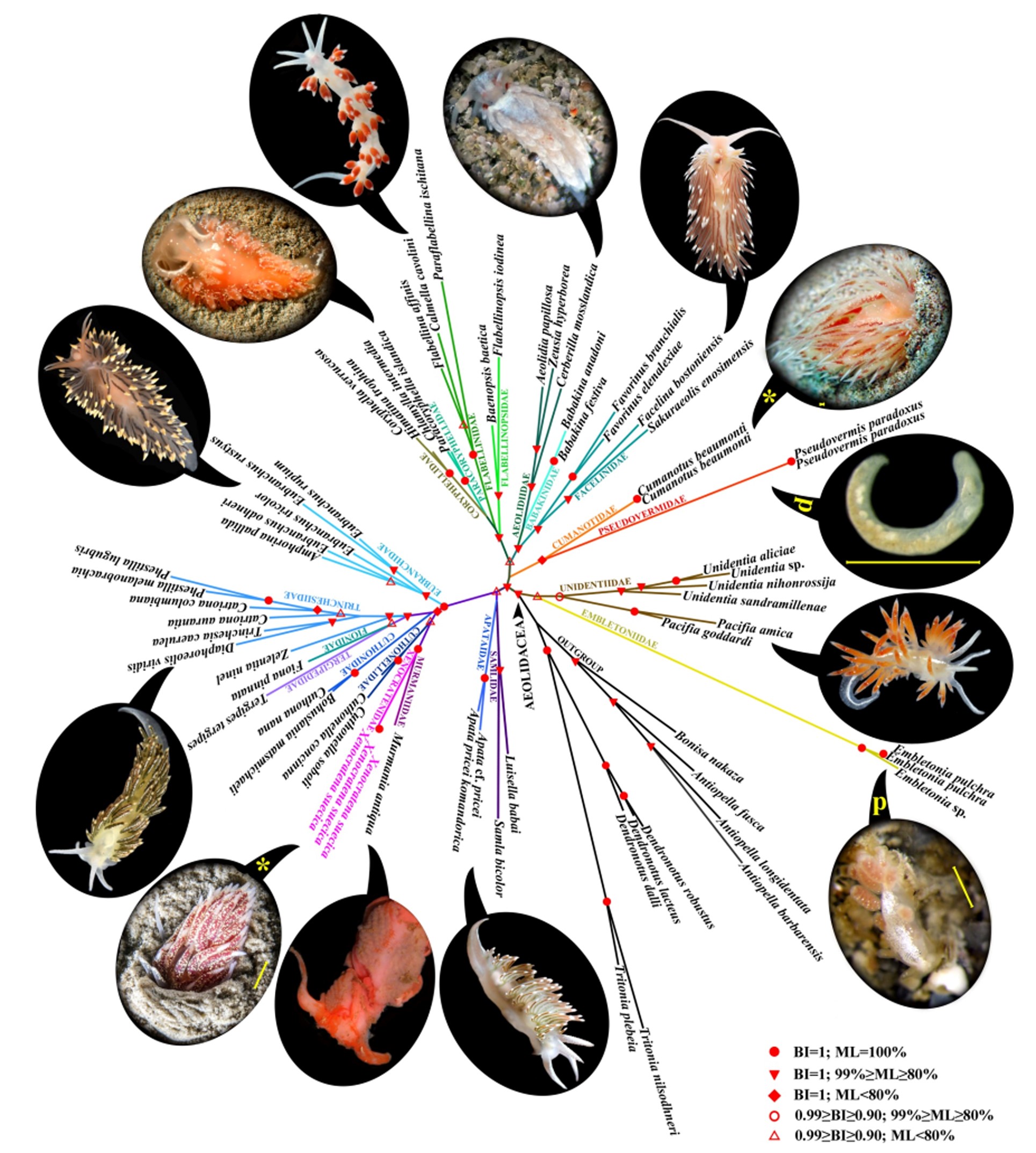
Рисунок 1. Молекулярно-филогенетический анализ Aeolidacea на основе пяти генетических маркеров (COI + 16S + 18S + 28S + H3).
Представитель Aeolidacea Xenocratena suecica был открыт в 1940 на мягких грунтах шведского Gullmar fjord. Повторных находок не было, и этот таксон оставался загадкой для исследователей вплоть до работы Martynov et al., 2020 (где помимо детально изученной морфологии, были получены первые молекулярные данные, рис. 1, 2). Молекулярно-филогенетический анализ выявил, что загадочная Xenocratena является сестринской группой семейства Murmaniidae, а род Xenocratena был выделен в самостоятельное семейство Xenоcratenidae (рис. 1). Одновременно, по числу и строению ветвей пищеварительной железы и соответствующих рядов спинных папилл, Xenocratena оказалась достоверно существенно проще, чем представители семейства Murmaniidae. Кроме того, взрослые Xenocratena, намного мельче Murmaniidae. Наконец, благодаря детальным подводным видео материалам, удалось продемонстрировать, что Xenocratena suecica целиком зарывается в мягкие грунты. Сочетание молекулярно-филогенетических и экологических данных показало, что Xenоcratenidae, являются педоморфными, упрощенными потомками более сложной предковой организации, но совершенно независимо от других педоморфных групп Aeolidacea, например, таких как Pseudovermidae. Таким образом, было достоверно показано, что обитание в мягких субстратах, несомненно, способствует вторичному упрощению и педоморфной эволюции.
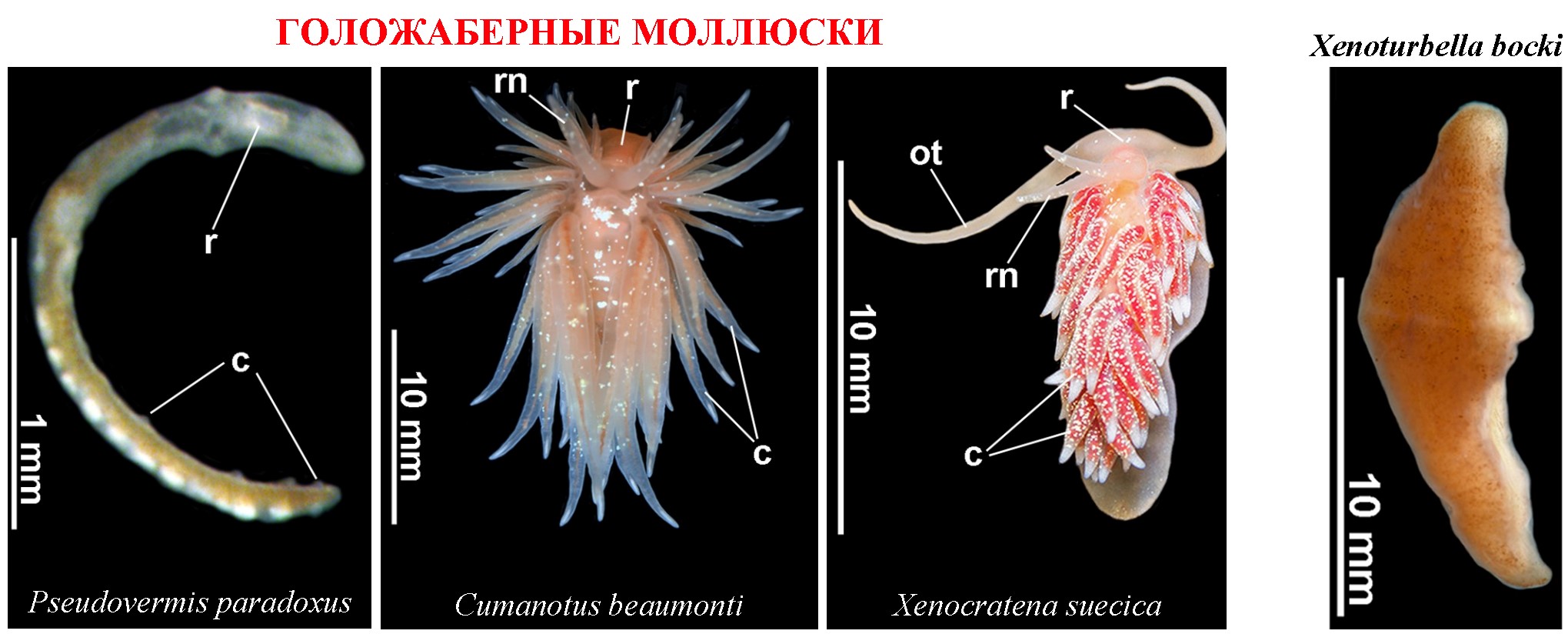
Рисунок 2. Pseudovermis paradoxus, Cumanotus beaumonti, Xenocratena suecica и Xenoturbella bocki, внешнее строение.
Интересно отметить, что в 1949 году (через 9 лет после открытия Xenocratena), из мягких грунтов того же Gullmar fjord было описано загадочное беспозвоночное животное Xenoturbella bocki (около 10 мм в длину, рис. 2). Позднее еще несколько представителей Xenoturbella были обнаружены на больших глубинах в Тихом океане. Организация Xenoturbella, на первый взгляд, очень проста: у них отсутствует сложная централизованная нервная система, нет целома и даже нефроцитов, а пищеварительный тракт слепо замкнут. Первоначально Xenoturbella стали относить к плоским червям. Проведенные впоследствии молекулярно-филогенетические исследования, тем не менее, показали куда более парадоксальную картину: во-первых, Xenoturbella оказалась сестринской группой бескишечных турбелярий Acoelomorpha, а во-вторых Xenoturbella + Acoelomorpha были выделены в отдельный тип билатерий Xenacoelomorpha, совершенно не родственный остальным плоским червям. Но на этом сюрпризы Xenoturbella не закончились. Тогда, как в ряде молекулярно-филогенетических работ, Xenacoelomorpha оказалась внешней группой по отношению ко всем остальным Bilateria (которых обозначили Nephrozoa – первичноротые + вторичноротые), в ряде других исследований было получено, что Xenoturbella +Acoelomorpha находятся среди настоящих билатерий, как сестринские, сложным иглокожим и полухордовым. Для группы, потенциально объединяющей Xenacoelomorpha, Echinodermata и “Hemichordata”, было предложено название Xenambulacraria. Таким образом, филогенетико-систематическое положение Xenoturbella и родственных ей бескишечных турбеллярий имеет непосредственное отношение к наиболее фундаментальным вопросам эволюции многоклеточных животных: в случае, если Xenoturbella является внешней группой к Bilateria, ее организацию можно рассматривать, как приближающуюся к таковой общего предка всех билатерий. Если же, на первый взгляд, очень простая Xenoturbella становится сестринской сложным иглокожим и полухордовым, то она является упрощенным потомком более сложных предковых Bilateria. В последнем случае, концепция “Nephrozoa” в основном теряет свое значение.
В работе Martynov et al., 2020 онтогенетико-систематическая модель была расширена на другие типы Bilateria, включая Xenoturbella: на эволюцию Xenoturbella могли повлиять те же факторы, которые подразумевают эволюцию от более сложных к упрощенным формам жизни, и было четко указано (в противовес широко поддерживаемому разными исследователями положению Xenacoelomorpha, как внешней группы по отношению ко всем Bilateria), что Xenoturbella могли происходить от сложных предков посредством педоморфной эволюции в мягких грунтах. То есть объяснение простого плана строения тела Xenoturbella заключается в том, что они произошли от более сложных урбилатерий за счет утраты морфологических признаков. На онтогонетико-систематической базе предложена модель происхождения cходных с Xenoturbella организмов от общего предкового онтогенетического цикла с типами Echinodermata, Pterobranchia и Enteropneusta, включая седентарную/полуседентарную взрослую организацию.
В 2024 году была опубликована работа большого международного исследовательского коллектива Schiffer et al., 2024, в которой был проанализирован геном Xenoturbella (~111 Mbp). Было выявлено, что Xenoturbella обладает сходным числом генов с другими билатериями, а также сохраняет предковую синтению Metazoa и ключевые сигнальные пути Bilateria, (что не соответствует мнимой простоте организации Xenoturbella). Например, количество генов Xenoturbella bocki (~15 000), Drosophila melanogaster (>14 000), C. elegans (~20 000). Длины интронов и экзонов также попадают в диапазон, наблюдаемый у многих других видов беспозвоночных. У Xenoturbella bocki выявили 31 нейропептидную систему, и для каждой найден, либо лиганд, либо рецептор, имеющиеся, как у первичноротых, так и у вторичноротых. Указывается, что геном Xenoturbella содержит почти полный набор нейропептидов билатерий без признаков упрощения (рис. 3). Несмотря на то, что Xenoturbella потеряла некоторые гены и часть компонентов сигнальных путей Bilateria, в целом геном Xenoturbella не является принципиально более простым, чем у других билатерий. Внимание также уделено возможному ошибочному влиянию на результаты молекулярно-филогенетических анализов, молекулярных данных Acoelomorpha (отличающихся высокими скоростями изменения последовательностей). В работе подчеркивается, что в результате проведенного анализа генома Xenoturbella, гипотеза Nephrozoa не получила поддержки.
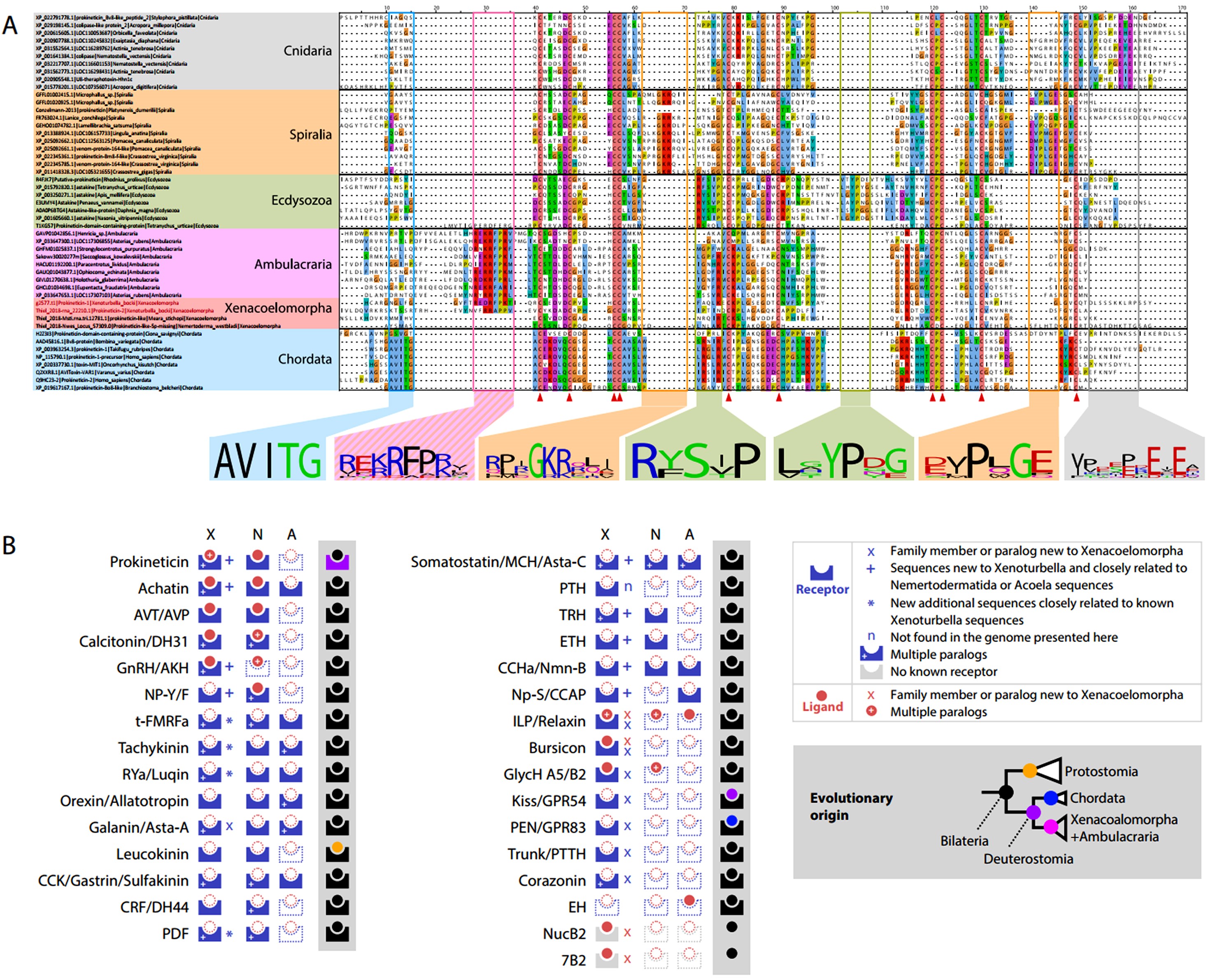
Рисунок 3. Геном Xenoturbella bocki содержит гены большинства билатерально-специфических пептидергических систем и ген прокинетицина, включающий signature последовательность, общую с Ambulacraria.
Новость подготовила
© Т.А. Коршунова
28.12.2024