 © Ильясов Рустем Абузарович, д-р биол. наук, ведущий научный сотрудник лаборатории нейробиологии развития.
© Ильясов Рустем Абузарович, д-р биол. наук, ведущий научный сотрудник лаборатории нейробиологии развития.
Научные интересы: апиология.
Пчелы (Anthophila) наиболее богатая видами клада из надсемейства Apoidea отряда перепончатокрылых Hymenoptera, содержащая более 20 000 видов, распределенных в семи семействах, которые распространены на всех обитаемых континентах. На сегодняшний день нет четкого понимания эволюционных процессов, которые привели к широкому современному распространению пчел и их роли в эволюции цветковых растений и формировании современных биоценозов (Ramirez et al., 2010; Bossert et al., 2022).
Согласно классической точке зрения, сформировавшейся на основе методов морфометрии и генетики, считается, что пчелы произошли в юго-восточной Азии, поскольку там наибольшее разнообразие видов пчел (Ilyasov et al., 2020). Однако вопрос о происхождении пчел остается до сих пор открытым и периодически появляются новые гипотезы. Альтернативную гипотезу о происхождении пчел излагает Michener (1979), по которой пчелы возникли в позднем меловом периоде в засушливых регионах Западной Гондваны (современная Южная Америка и Африка) (Michener, 1979). В гипотезе Michener было много разногласий с современными молекулярными данными, которые не позволяли признать ее верной. Однако международная группа исследователей Almeida et al. (2023) на основе геномных и ископаемых данных с применением современных статистических и биоинформационных методов анализа подтвердила верность гипотезы Michener (1979). В статье в журнале Current Biology (2023) Almeida et al. представили новую реконструкцию исторической биогеографии пчел, показали их происхождение в Западной Гондване в раннем меле, одновременно с первыми цветковыми растениями около 120 миллионов лет назад. На основе объединенных филогеномных и палеобиогеографических исследований Almeida et al. (2023) выполнили подробную таксономическую реконструкцию, включающую 216 современных видов из всех признанных в настоящее время 7 семейств и 28 подсемейств пчел, представленных во всех биогеографических регионах мира (рис. 1).
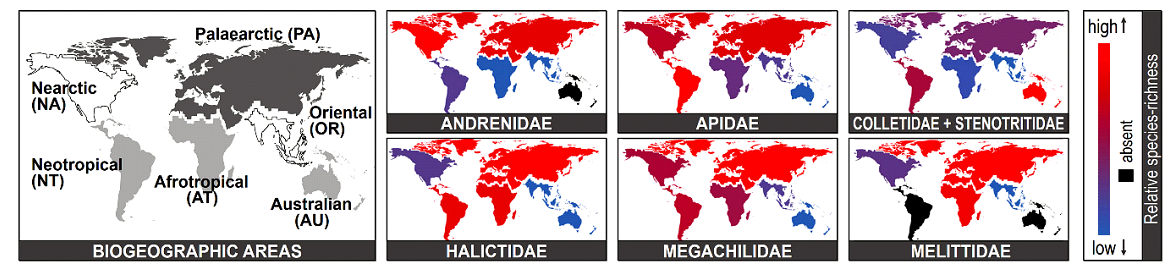
Рис. 1. Современное географическое распределение семи крупных семейств пчел в разных биогеографических регионах мира. Красный цвет - высокое видовое разнообразие, синий цвет - низкое видовое разнообразие, черный цвет - отсутствуют представители (Almeida et al., 2023).
На основе оценки 185 ископаемых видов пчел была реконструирована временная шкала дивергенции пчел. Данная реконструкция эволюции пчел является на сегодня самой масштабной работой с доказательствами из палеонтологических и геномных исследований. На филогенетическом древе представлены все семейства и подсемейства пчел (рис. 2) (Almeida et al., 2023).
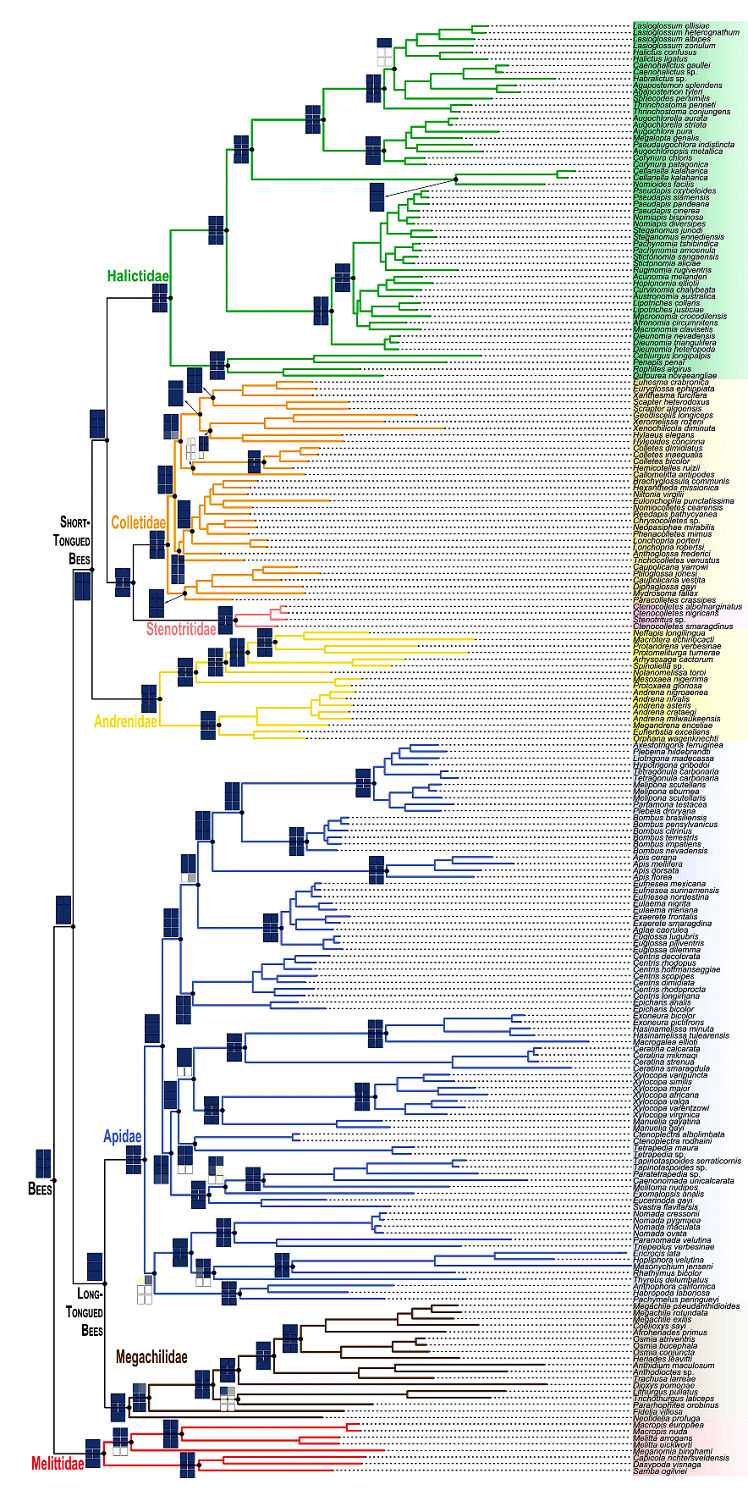
Рис. 2. Реконструкция филогенетики пчел (Hymenoptera: Apoidea: Apiformes), выведенная на основе анализа 830 локусов ультраконсервативных элементов (UCE) и общей гетерогенной эволюции на модели единой топологии (GHOST), построенная в IQ-TREE (Almeida et al., 2023).
Согласно Almeida et al. (2023), пчелы возникли в раннем меловом периоде (106,6-137,9 млн лет назад, среднее значение: 124 млн лет назад). Все расхождения между основными семействами пчел произошли между средним и поздним меловым периодом, около 73–124 млн лет назад. Предполагается, что все основные современные семейства пчел дивергировали и дифференцировались до начала третичного периода (Engel, 2001). Результаты Almeida et al. (2023) контрастируют с другими исследованиями, в которых пчелы имели более молодой возраст, что, вероятно, связано с ограниченной выборкой таксонов пчел, методологическими различиями в калибровке времени происхождения ископаемых пчел (рис. 3) (Branstetter et al., 2017; Sann et al., 2018; Almeida et al., 2023).
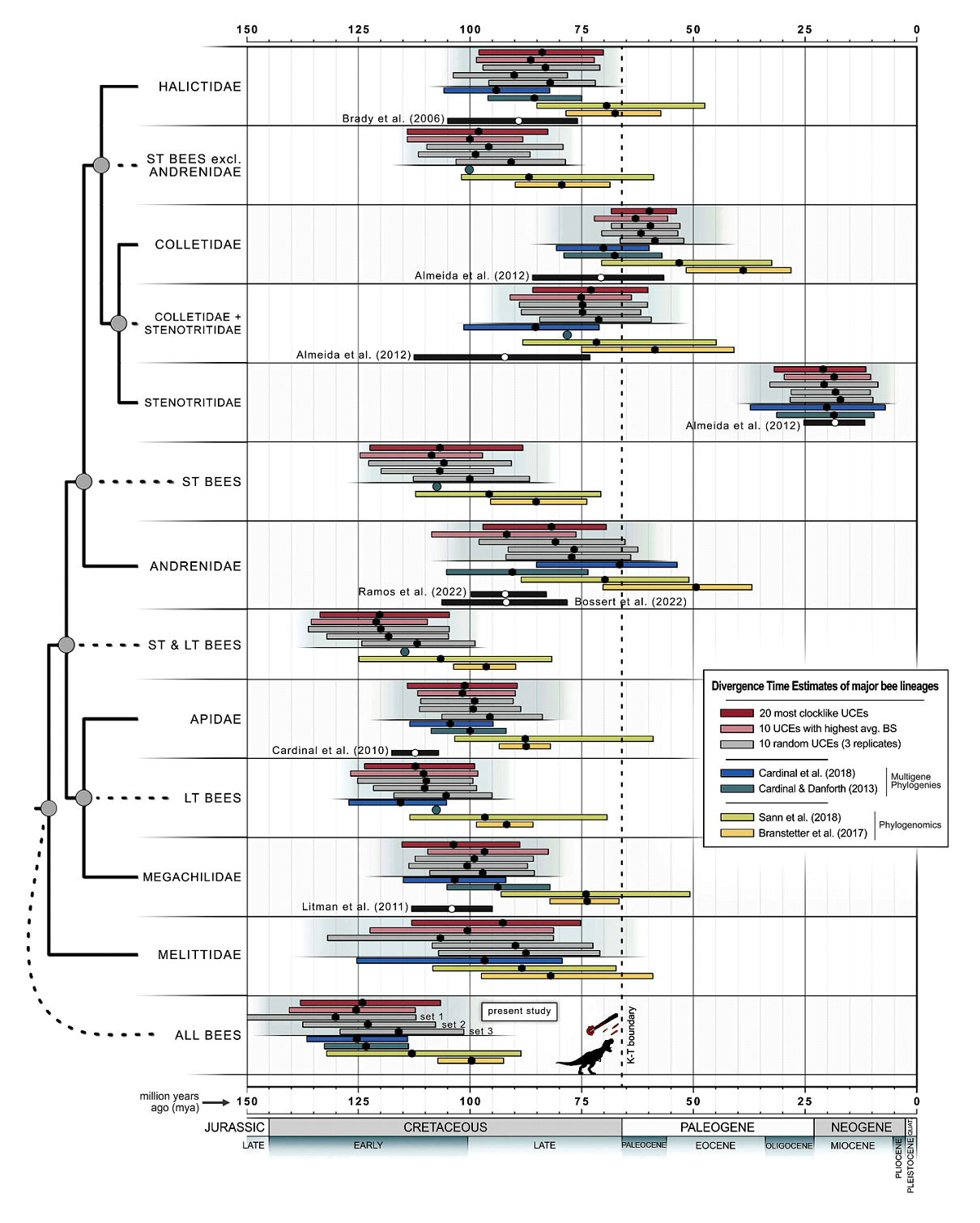
Рис. 3. Время дивергенции 13 семейств пчел (Apoidea: Apiformes). Сравнение результатов геномных и палеонтологических исследований Almeida et al. (2023) с результатами других исследователей. Дендрограмма слева отражает филогению пчел на уровне семейств; ST - пчелы с коротким язычком (Andrenidae, Colletidae, Halictidae и Stenotritidae), а LT - пчелы с длинным язычком (Apidae и Megachilidae) (Almeida et al., 2023).
Разделение Южной Америки и Африки примерно 100 млн лет назад привело к разделению двух семейств пчел с длинным язычком (Apidae и Megachilidae) на африканские и южноамериканские линии. Согласно их анализу, Melittidae оставались ограниченными Африкой либо потому, что они были пространственно ограничены восточной частью Западной Гондваны, либо потому, что линии Melittidae в Южной Америке впоследствии вымерли. Это делает Melittidae единственным существующим семейством пчел, которое, вероятно, претерпело раннюю диверсификацию исключительно в Восточном полушарии. Тесная связь Melittidae с Африкой также согласуется с наблюдением, что это единственный континент, на котором обитают все три подсемейства (Danforth et al., 2006). Тектонические события мелового периода также ограничили предков современных Colletidae, Andrenidae и Halictidae Южной Америкой. Некоторые специализированные ассоциации между пчелами и опыляемыми растениями, вероятно, возникли в меловом периоде. Возможно, повышенное разнообразие цветковых растений в тропической зоне связано с их длительной коэволюцией с пчелами (Hughes et al., 2013). Филогенетические реконструкции предполагают, что в середине мелового периода пчелы еще не достигли Австралии и Индийского субконтинента - там не было пчел вплоть до эоцена, пока они оставались изолированным от других континентов. Это согласуется с наблюдением, что не существует древних эндемичных линий пчел на территории Индии (Almeida et al., 2023).
Все существующие семейства пчел, вероятно, отделились друг от друга к концу мелового периода, 80 млн лет назад, и распределились по всем трем южным континентам. Это можно объяснить первым расширением ареала на Австралийский континент. К позднему меловому периоду из Южной Америки в Австралию проникли семейства Colletidae и Stenotritidae. Этот переход произошел с мелового периода до эоцена, когда Южная Америка, Антарктида и Австралия были связаны и обладали сходным климатом. В настоящее время 50% австралийской фауны пчел составляют Colletidae и Stenotritidae. Исследования эндемичных австралийских групп Colletidae, таких древних австралийских подсемейств как Euryglossinae, Hylaeinae и Neopasiphaeinae, показывают, что эти линии сохраняют узкую ассоциацию с преобладающими в Австралии семействами цветковых растений Myrtaceae, Fabaceae и Proteaceae. Другие линии пчел, присутствующие в настоящее время в Австралии (Apidae, Megachilidae и Halictidae), прибыли в Австралию намного позже (рис. 4) (Almeida et al., 2023). Согласно реконструкциям по ископаемым образцам Almeida et al. (2023), пчелы Apidae и Megachilidae начали колонизировать Северное полушарие в позднем меловом периоде и раннем палеогене (60–40 млн лет назад) через связи между Южной и Северной Америкой, а также между Африкой и Европой. Было предложено шесть биогеографических схем кладогенеза пчел в результате расширения и сокращения ареалов. Биогеографическая интерпретация существующих и ископаемых пчел представлена на палеокартах, представляющих геологические реконструкции от 120 млн лет назад до наших дней с использованием байесовского анализа кладогенеза в результате расширения и сокращения ареалов ископаемых групп пчел (рис. 4) (Almeida et al., 2023).
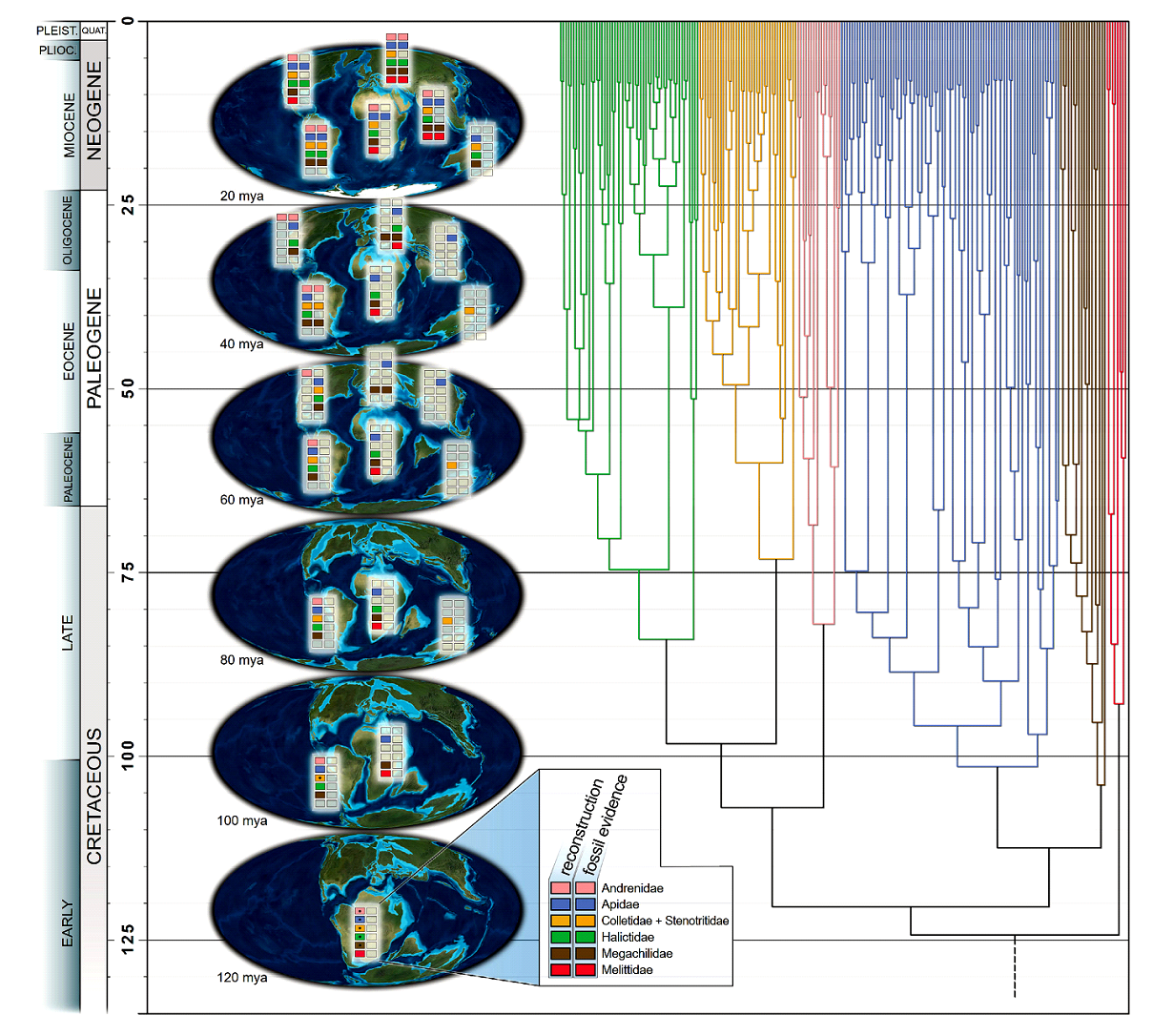
Рис. 4. Биогеографическая реконструкция эволюции пчел на основе ископаемых и современных модельных аналитических данных. Цветные прямоугольники указывают на предполагаемое присутствие семейств пчел в регионе (Almeida et al., 2023).
Палеоцен и эоцен отмечались тепловыми максимумами, когда тропический и субтропический климат охватывал более высокие широты. Эти периоды были климатически благоприятными для таксонов пчел, ограниченных теплыми условиями, что, вероятно, позволило им достичь и временно расширить свои ареалы на север. В палеоцен-эоценовые периоды происходило расширение ареалов пчел Andrenidae и Apidae на север (Bossert et al., 2022; Freitas et al., 2022). Некоторые таксоны (Anthophorinae) впоследствии вымерли в более высоких широтах, вероятно, когда климат перестал быть благоприятным (Orr et al., 2022). Лучшим примером таких пространственно-временных изменений ареалов являются безжальные пчелы (Apidae: Meliponini). В настоящее время они обитают только в тропических регионах, но их ископаемые представители, например, Cretotrigona prisca найдены в районах Северной Америки и Европы, что может служить доказательством их прошлых экспансий на север (Michener and Grimaldi, 1988; Engel, 2000; Almeida et al., 2023).
В поздний палеоген, эоцен, олигоцен (40 млн лет назад) и неоген (20 млн лет назад) различные таксоны пчел продолжали мигрировать между Южной и Северной Америкой, Африкой и Европой, а также между Евразией и Северной Америкой через Берингов и Североатлантический сухопутные мосты. Melittidae, ареал которых ранее был ограничен Африкой, распространились к северу от Африки. Группы, диверсифицировавшиеся на южных континентах после периодов эоценового глобального потепления и закрытия бореотропического коридора (Oxaeinae, Protandrenini, Emphorini), не расширили свое распространение так сильно, как таксоны, которые диверсифицировались раньше. Глобальные вымирания, частично наблюдаемые с помощью ископаемых пчел, также ознаменовали поздний эоцен, например, вымирание корбикулятных пчел Melikertini (Engel, M.S. 2001; Almeida et al., 2023)
Согласно реконструкции биогеографии пчел Almeida et al. (2023), все семейства пчел достигли Южной Азии в неогене или немного ранее (рис. 3). Вероятно, Apidae, Megachilidae, Halictidae достигли Австралии через Южную Азию и Новую Гвинею. Вплоть до позднего олигоцена - неогена фауна пчел Австралии целиком состояла из двух близкородственных семейств (Colletidae и Stenotritidae). Вероятно, ранняя эволюция флоры цветковых растений в Индии происходила в отсутствие пчел (Scotese, 2004; Seton et al., 2012). Пчелы в Индии появились около 50 млн лет назад после ее столкновения с Азией согласно найденным там ископаемым данным вымерших линий пчел Apidae из раннего эоцена (Rutschmann et al., 2004; Sanil et al., 2022). Данное событие сильно повлияло на местное разнообразие флоры Индии (Almeida et al., 2023).
Таким образом, на основе большого объема геномных и ископаемых данных Almeida et al. (2023) показали, что пчелы возникли в раннем меловом периоде в западной части Гондваны (Африка и Южная Америка) около 120 миллионов лет назад. На их биогеографическую историю впоследствии повлияла фрагментация Гондваны (разделение пчел южноамериканских и африканских линий) и расширение географического распространения в Северном полушарии. Пчелы колонизировали северные континенты благодаря существованию мостов и благоприятному климату в исторический период. Раннее отделение Индии от других южных континентов привело к тому, что Индия была «свободной от пчел» на протяжении значительной части своей геологической истории. Точно так же в Австралии, которая также была изолирована на раннем этапе, на протяжении большей части ее геологической истории не было пчел и только с позднего мелового периода туда проникли пчелы двух близкородственных семейств (Colletidae и Stenotritidae). Дополнительным подтверждением происхождения пчел в Южном полушарии может являться высокое таксономическое разнообразие современных цветковых растений в тропических регионах. Раннее отсутствие пчел на больших массивах суши, особенно в Австралии и Индии, имеет важное значение для понимания коэволюции местной флоры и пчел опылителей. Реконструкция эволюции пчел дает новое представление о влиянии пчел посредством опыления на филогенез и таксономическое разнообразие цветковых растений с мелового периода (Almeida et al., 2023).
Список литературы- Almeida, E.a.B., Bossert, S., Danforth, B.N., Porto, D.S., Freitas, F.V., Davis, C.C., Murray, E.A., Blaimer, B.B., Spasojevic, T., Stroher, P.R., Orr, M.C., Packer, L., Brady, S.G., Kuhlmann, M., Branstetter, M.G., Pie, M.R., 2023. The evolutionary history of bees in time and space. Current Biology. doi: 10.1016/j.cub.2023.07.005.
- Bossert, S., Wood, T.J., Patiny, S., Michez, D., Almeida, E.a.B., Minckley, R.L., Packer, L., Neff, J.L., Copeland, R.S., Straka, J., Pauly, A., Griswold, T., Brady, S.G., Danforth, B.N., Murray, E.A., 2021. Phylogeny, biogeography and diversification of the mining bee family Andrenidae. Systematic Entomology. 47 (2), 283-302. doi: 10.1111/syen.12530.
- Branstetter, M.G., Danforth, B.N., Pitts, J.P., Faircloth, B.C., Ward, P.S., Buffington, M.L., Gates, M.W., Kula, R.R., Brady, S.G., 2017. Phylogenomic Insights into the Evolution of Stinging Wasps and the Origins of Ants and Bees. Curr Biol. 27 (7), 1019-1025. doi: 10.1016/j.cub.2017.03.027.
- Danforth, B.N., Sipes, S., Fang, J., Brady, S.G., 2006. The history of early bee diversification based on five genes plus morphology. Proceedings of the National Academy of Sciences of the United States of America. 103 (41), 15118-23. doi: 10.1073/pnas.0604033103.
- Engel, M.S., 2000. A new interpretation of the oldest fossil bee (Hymenoptera, Apidae). American Museum Novitates. 3296, 1–11.
- Engel, M.S., 2001. Monophyly and extensive extinction of advanced eusocial bees: insights from an unexpected Eocene diversity. Proc Natl Acad Sci U S A. 98 (4), 1661-4. doi: 10.1073/pnas.98.4.1661.
- Hughes, C.E., Pennington, R.T., Antonelli, A., 2013. Neotropical Plant Evolution: Assembling the Big Picture. Botanical Journal of the Linnean Society. 171 (1), 1-18. doi: 10.1111/boj.12006.
- Ilyasov, R.A., Lee, M.-L., Takahashi, J.-I., Kwon, H.W., Nikolenko, A.G., 2020. A revision of subspecies structure of western honey bee Apis mellifera. Saudi Journal of Biological Sciences. 27 (12), 3615-3621. doi: 10.1016/j.sjbs.2020.08.001.
- Michener, C.D., 1979. Biogeography of the Bees. Annals of the Missouri Botanical Garden. 66 (3), 277. doi: 10.2307/2398833.
- Michener, C.D., Grimaldi, D.A., 1988. A Trigona from Late Cretaceous amber of New Jersey (Hymenoptera: Apidae: Meliponinae). American Museum Novitates. 2917, 1–10.
- Ramírez, S.R., Roubik, D.W., Skov, C., Pierce, N.E., 2010. Phylogeny, diversification patterns and historical biogeography of euglossine orchid bees (Hymenoptera: Apidae). Biological Journal of the Linnean Society. 100, 552-572. doi: 10.1111/j.1095-8312.2010.01440.x.
- Rutschmann, F., Eriksson, T., Schönenberger, J., Conti, E., 2004. Did Crypteroniaceae Really Disperse Out of India? Molecular Dating Evidence fromrbcL,ndhF, andrpl16 Intron Sequences. International Journal of Plant Sciences. 165 (S4), S69-S83. doi: 10.1086/383335.
- Sanil, M.S., Balakrishnan, S., Sreekumar, V.B., Dev, S.A., 2022. Dipterocarps used India as a raft from Gondwana to Eurasia. Taxon. 71 (6), 1214-1229. doi: 10.1002/tax.12794.
- Sann, M., Niehuis, O., Peters, R.S., Mayer, C., Kozlov, A., Podsiadlowski, L., Bank, S., Meusemann, K., Misof, B., Bleidorn, C., Ohl, M., 2018. Phylogenomic analysis of Apoidea sheds new light on the sister group of bees. BMC evolutionary biology. 18 (1), 71. doi: 10.1186/s12862-018-1155-8.
- Scotese, Christopher r., 2004. A Continental Drift Flipbook. The Journal of Geology. 112 (6), 729-741. doi: 10.1086/424867.
- Seton, M., Müller, R.D., Zahirovic, S., Gaina, C., Torsvik, T., Shephard, G., Talsma, A., Gurnis, M., Turner, M., Maus, S., Chandler, M., 2012. Global continental and ocean basin reconstructions since 200Ma. Earth-Science Reviews. 113 (3-4), 212-270. doi: 10.1016/j.earscirev.2012.03.002.
Новость подготовил
© Ильясов Р.А.
28.08.2023