 © Симонова Ольга Борисовна
− руководитель лаборатории молекулярно-генетических процессов развития, доктор биологических наук.
© Симонова Ольга Борисовна
− руководитель лаборатории молекулярно-генетических процессов развития, доктор биологических наук.
Исследователи из Института наук о жизни при Университете Мичигана (University of Michigan Life Sciences Institute) и Медицинского института Говарда Хьюза (Howard Hughes Medical Institute) определили, что сателлитная ДНК прицентромерных районов, которая (в отличие от ДНК центромерных районов) долго считалась "мусорной ДНК" ("junk DNA"), играет важнейшую роль в сборке генома. Выводы ученых, опубликованные недавно в журнале eLife, говорят о том, что генетический «мусор» прицентромерных районов играет жизненно важную функцию, обеспечивая локализацию хромосом в пределах ядра клетки, что необходимо для её выживания. И эта функция, по всей видимости, является эволюционно консервативной.
Перицентрическая сателлитная ДНК состоит из очень простой и часто повторяющейся последовательности нуклеотидов. И хотя на ее долю приходится значительная часть нашего генома, сателлитная ДНК не содержит каких-либо конкретных инструкций по созданию каких-либо конкретных белков, т.е. является некодирующей. Какое-то время ученые считали, что «мусорная», или «эгоистичная», ДНК перицентрических районов не играет никакой роли и не служит никакой цели, поскольку последовательности повторяющихся нуклеотидов этих районов сильно вариабельны и не являются консервативными даже у близкородственных видов. Ямашита, профессор Мичиганского института Life Sciences Institute, и ее коллеги решили посмотреть, что произойдет, если клетки не будут использовать перицентромерную сателлитную ДНК.
Поскольку сателлитная ДНК существует в виде длинных повторяющихся последовательностей, ученые не могли просто мутировать или вырезать всю эту ДНК из генома. Вместо этого они использовали нулевой аллель гена D1, продукт которого, белок D1, связывается с сателлитной ДНК в АТ-богатых районах гетерохроматина у D. melanogaster. Сначала они показали, что белок D1 дрозофилы, как и его гомолог у мыши, важны для формирования особой структуры хромосом, которая объединяет их перицентрические районы в единый компартмент – хромоцентр (рис. 1). Отметим, что хромоцентр был открыт ещё 50 лет назад, однако, мало кто исследовал его функции за рубежом. Затем, используя дрозофил с делецией гена D1, ученые удалили D1 из клеток плодовой мушки и тут же обнаружили, что зародышевые клетки полового пути – предшественники сперматозоидов или яйцеклеток – погибают. Дальнейший анализ показал, что в этих клетках идет нарушение формирования хромоцентра, и на стадии интерфазы формируются крошечные, отпочковывающиеся за пределы ядра, микроструктуры несущие части генома, которые затем превращаются в микроядра (рис. 2). Без всего генома, инкапсулированного в ядро, клетки не могли выжить.
Ученые считают, что белок D1 связывается с сателлитной ДНК, собирая все хромосомы в ядре, формируя хромоцентр. Если белок D1 не может связаться с сателлитной ДНК, клетка теряет способность образовывать полноценное ядро и погибает. Исследователи провели аналогичные эксперименты с использованием клеток мышей и получили те же результаты. Когда они удаляли гомологичный белку D1 белок HMGA1 мыши, который также связывается с прицентромерной ДНК, клетки снова образовывали микроядра и не выживали.
Полученные в экспериментах на дрозофиле и мышах данные заставляют команду полагать, что сателлитная прицентромерная ДНК необходима для выживания клеток, имеющих ядра, у разных видов, включая человека, а функция хромоцентра эволюционно консервативна.
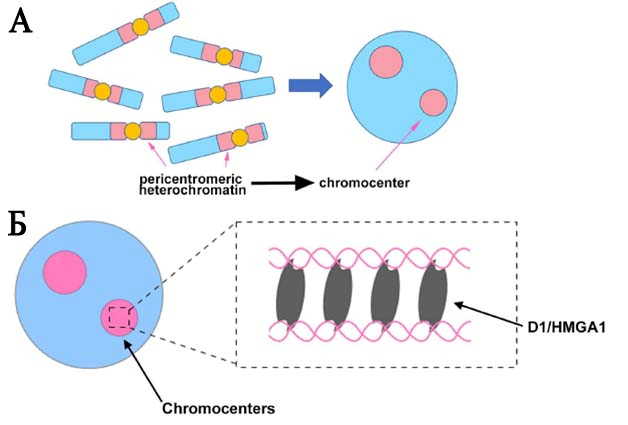
Рисунок 1.
А. Области перицентромерных районов хромосом (розовый цвет слева) формируют хромоцентр (розовые круги справа). Район центромерного хроматина показан желтым цветом.
Б. Модель функционирования белка D1 дрозофилы и гомологичного белка HMGA1 мыши. Показано, как эти белки связываются с ДНК гетерологичных хромосом, объединяя их в хромоцентр (розовый круг).
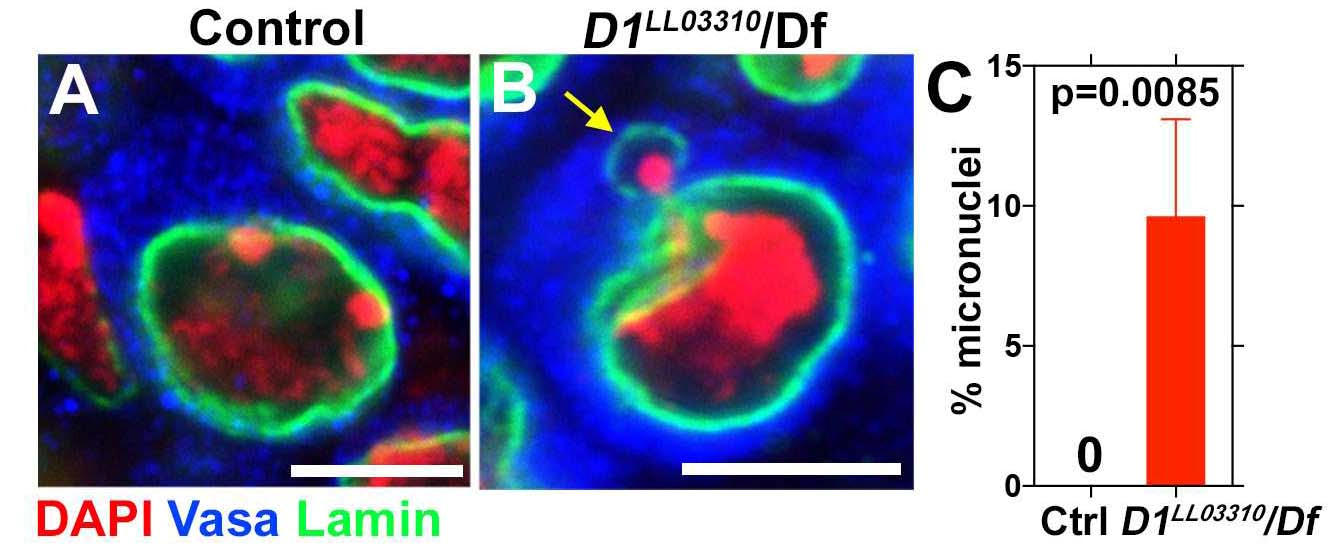
Рисунок 2.
Мутация гена D1 у дрозофилы и РНК-и опосредованное ингибирование HMGA1 мыши увеличивает частоту формирования микроядер.
А – нормальные клетки полового пути дрозофилы;
В – клетки полового пути мутанта с делецией гена D1,
жёлтая стрелка указывает на почкующееся микроядро;
C – Частота формирования микроядер в контроле (Ctrl) и у мутанта (D1/Df).
DAPI – окраска ДНК,
Vasa – окраска цитоплазмы герминативных (половых) клеток,
Lamin – окраска ядерной оболочки.
Новость подготовила © 2017 Симонова О.Б.
28.04.2018