 © Чистопольский Илья Александрович, к.б.н., инженер лаборатории нейробиологии развития ИБР.
© Чистопольский Илья Александрович, к.б.н., инженер лаборатории нейробиологии развития ИБР.
Область научных интересов: нейробиология, биология поведения.
Статья «The Mechanical Basis of Memory – the MeshCODE Theory» посвящена одной из базовых проблем биологии. В ней представлена новая гипотеза о локализации памяти в измененной конформации белков при работе мозга млекопитающего. Делается попытка логически согласовать работу предполагаемого механизма с известными принципами работы мозга.
Подход автора имеет свою историю. Здесь уместно сослаться на Пенроуза (R.Penrose, 1998), который, исходя из философских и логических соображений и анализируя математическое мышление человека, полагает, что сознательная деятельности мозга должна опираться на элементы, имеющие существенно меньший размер, чем отдельная клетка. Принципиальная неалгоритмичность некоторых математических методов, которые применяет человек для решения математических задач, заставила Пенроуза сделать вывод о том, что работа сознания связана с процессами в мозге на уровне микромасштабов. Т.е. это масштаб отдельных молекул, на котором непригодны к использованию термодинамические законы и необходимы к применению законы квантовой физики. Умозрительный вывод Пенроуза сходен с тем, к которому пришли и некоторые биологи. Они полагают, что базовые процессы мышления, реализуемые в нервной ткани, опираются на структуры значительно меньших размеров, чем нейрон, и расположены внутри него. Ключевую роль, с их точки зрения, здесь играют те или иные филаменты. Например, Хамерофф (Hameroff S.R., 1996) полагал, что ключевую роль тут играют микротрубочки из тубулина. Еще одна гипотеза, сходная с этой, находит основу мыслительных процессов не в самом тубулине, а в воде внутри микротрубочек. В этом случае, вода вблизи белка структурирована на масштабах в несколько нм и её поведение определяется законами квантовой физики. Другой исследователь, Либерман Е.А. (1995), предполагал, что при работе нейрона становится критична энергия затрат на обработку информации и связывал основную «вычислительную» работу нейрона с механическими филаментами внутри нейрона. Это позволяло ему ожидать большей энергетической эффективности от работы каждого отдельного нейрона.
Все перечисленные выше гипотезы предполагают распространение сигналов по упругим филаментам тем или иным способом и взаимодействие этих филаментов с белками мембран нейронов как со стороны входов (постсинапсы дендритов) так и со стороны аксона (пресинапсы терминалей). Описанные выше подходы очень похожи на гипотезу автора статьи. Вместе с тем, в них уделялось мало внимания памяти, что значительно сужало применимость моделей. Автор статьи сделал следующий шаг вслед за указанными выше «предшественниками».
Основную идею он формулирует так:
«… память хранится в мозге в механически закодированном двоичном формате, записанном в конформациях белков, обнаруженных в соединениях (adhesions) межклеточного матрикса (ECM ) cell-extracellular matrix, которые организуют каждый синапс. Описанная здесь структура MeshCODE представляет собой объединяющую теорию хранения данных у животных, обеспечивая хранение как динамической, так и постоянной информации в двоичном формате для чтения и записи. Механочувствительные белки, содержащие зависимые от силы переключатели, могут постоянно хранить информацию, которая может быть записана или обновлена под действием слабых механических сил.
Эти механочувствительные белки, такие как talin, образуют каркас каждого синапса, создавая сеть (meshwork) переключателей, которые вместе образуют код, так называемый MeshCODE. Большие сигнальные комплексы собираются на этих каркасах в зависимости от паттернов переключения, и эти комплексы должны стабилизировать паттерны и координировать синапсы, чтобы динамически настраивать синаптическую активность. Последовательности импульсов синаптической передачи и потенциалов действия будут управлять цитоскелетным аппаратом для записи и обновления синаптических MeshCODE…»
Классическая электрохимическая синаптическая передача в этой схеме служит только для активации считывания и перезаписи кода информации, который, по сути, и является памятью, а сам код и «машина» для его изменения реализуется белками цитоскелета каждого нейрона и белками межклеточного матрикса (ЕСМ).
Автор выдвигает предположение что основным белком, который служит материалом для хранения информации, является talin. Его свойство по сохранению нескольких стабильных конформаций позволяет полагать его как элемент-переключатель, который, при определенных условиях, может сохранять каждое из своих метастабильных состояний достаточно долго (см. рис.1 из статьи)
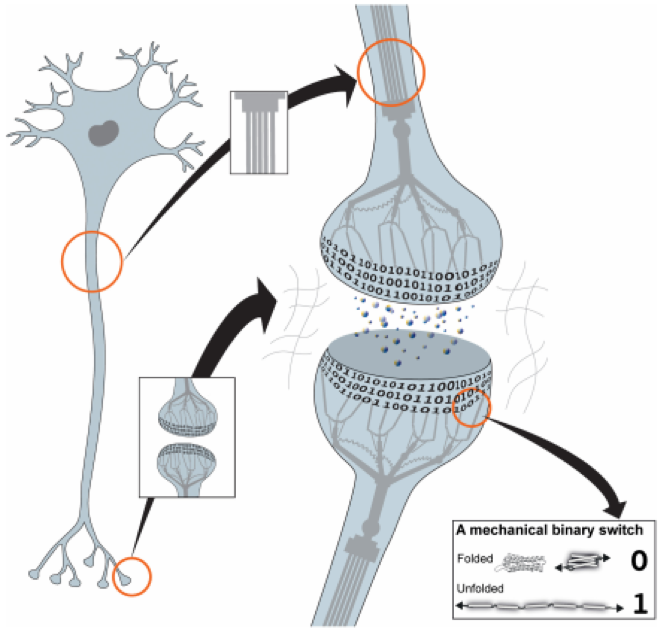
Автор полагает, что все белки цитоскелета клетки, а также белки межклеточной адгезии вокруг клеток, ЕСМ, связаны в единую сложную систему сети. Т.е. все процессы, связанные с передачей информации нейронов (синтез белка, филаменты, мембраны и везикулы синапсов и т.п.), интегрируются с помощью этой сети. Устойчивое состояние (гомеостаз) локальной сети каждого нейрона не единственно. Выбор конкретного устойчивого состояния сети однозначно определяется внешними и внутренними факторами для каждого нейрона. Вся регулировка состояний сводится к механическим сдвигам в этой общей сети, в следствие которых наступает новое термодинамическое равновесие сети. В этом новом состоянии часть доменов talin-а, локализованные в синапсах и встроенные в сеть, меняют свою конформацию и тем самым маркируют вновь наступившее равновесие. Такая жесткая с точки зрения устойчивости состояний сеть, внутри нейронов и проходящая сквозь них, позволяет избегать потерь данных в случае механических сдвигов внутри тканей, т.к. внутриклеточная и внеклеточная среды служат хорошей защитой ЕСМ от внешних деформаций.
Фиксирование состояния сети на уровне отдельного нейрона, например, в процессах пластичности при обучении, будет проявлять себя в регулировке цитоскелета при выбросе везикул, в конкуренции встраивания вновь синтезированного белка в более активных синапсах, в регулировке белков пре и пост синаптических мембран, регулировке экспрессии генов вплоть до эпигенетических модификаций. По замыслу автора это все, в целом, позволяет регулировать электрохимические соединения нейронов, опираясь на прошитую в памяти нейрональной ткани информацию, а память задается конформацией доменов talin-a и, в конечном итоге, всей механической сетью ЕСМ.
По данным автора, фиксация перестройки ЕСМ при смене конформаций talin-a происходит в течении нескольких минут, т.е. в это время другие белки-лиганды «закрепляют» происходящие деформации, а также деформации самого talin-a. Это позволяет автору выделить класс неустойчивой памяти без фиксации, быструю временную память. Но наибольший интерес для автора представляет возможность длительной фиксации состояния сети интегринами ЕСМ, т.е. стабилизации новых состояний сети после механических сдвигов при «перекодировании» белка talin-а в синапсах. Основная работа статьи – детальное описание возможных механизмов изменения белков синапсов нейрона и последующая реализация этих белковых изменений как памятного следа в мозге.
Автор полагает, что в его гипотетической системе используется двоичное кодирование, что, в общем случае, не обязательно. Вместе с тем, это дает ему возможность использовать аналогии с современной компьютерной техникой. Автор предполагает, что кодирование связывает работу многих отдельных частей мозга в единое целое, одновременно сохраняя рассредоточенный вид хранения памяти по многим нейронам. Следуя за компьютерной аналогией, он формулирует ряд гипотез. Он полагает, что колончатые структуры неокортекса мозга представляют собой удобные элементы памяти, а известная на сегодня активная работа мозга в течении фазы «быстрого» сна представляет собой процесс перезаписи памяти в неокортекс из гиппокампа с использованием молекулярного механизма механических изменений ЕСМ. Компьютерная аналогия приводит автора к тому, что гиппокамп необходим для правильной адресации информации. Процессы записи и перезаписи в этом случае идут посредством точного кодирования в сети ЕСМ каждого нейрона и такого рода активность может быть не связана с классической активностью нейронов, т.е. с их обычными потенциалами действия.
Последние гипотезы автора о работе целого мозга необходимо предполагают точное и дискретное кодирование. Если возможная стохастичность организации синапса прямо не противоречит заявленному механизму работы жестких ЕСМ сетей, то детерменизм в адресации записи памяти, предполагает, что любая стохастичность архитектуры нейрональных связей вызовет только зашумление цепей передачи полезных сигналов. Таким образом, в представленной гипотезе автора нет места случайно распределенным элементам, что в настоящее время выглядит умозрительным. Вместе с тем вопрос допустимой меры случайности при организации архитектуры отдельных сетей ЕСМ в статье не обсуждается.
Его подход заставляет поставить и еще один вопрос: А как долго без деградации способны существовать конструкции ECM и talin-а? В исследованиях памяти долговечность субстрата-носителя памятного следа – основной вопрос, который не имеет очевидного ответа в настоящее время. Например, Кандель (E. Kandel, 2006), в попытке ответить на него, одно время полагал, что в синапсах возможны белки прионовой природы. Современные представления, локализуя основную долговременную память в синапсах, рассматривают её стабилизацию или (а) через стабильность молекул матрикса синапса или (б) через устойчивость уровня синтеза молекул синапса. Оба подхода имеют как недостатки, так и преимущества. В модели автора дискретность и точность «хранения» информации в синапсах имеют принципиальное значение, но долговременная стабильность молекул, которые формируют в синапсе памятный след, в статье не обсуждается.
Заявленная автором гипотеза позволяет по-новому взглянуть на классическую проблему синаптической пластичности и памяти. Детальность формулировок представленной гипотезы даёт возможность её экспериментальной проверки в дальнейшем. Вместе с тем в статье, в основном, рассматриваются информационные аспекты модели, а принципиальные моменты биологии модели остаются за рамками обсуждения.
Новость подготовил © Чистопольский И.А.
27.05.2021