 © Баклушинская Ирина Юрьевна, ведущий научный сотрудник лаборатории эволюционной генетики развития, доктор биологических наук.
© Баклушинская Ирина Юрьевна, ведущий научный сотрудник лаборатории эволюционной генетики развития, доктор биологических наук.
Трудно переоценить значение мейоза, процесса, который способствует увеличению генетического разнообразия благодаря рекомбинации и обеспечивает стабильность генома вида. Один из подходов при изучении природных явлений - анализ отклонений, исключений из правил. Слепушонки - единственная группа млекопитающих, для которых в норме характерны морфологически идентичные половые хромосомы XX и у самцов, и у самок. Выбор адекватной модели нередко определяет успех исследования. В этом отношении слепушонки (рис. 1), небольшие грызуны, живущие под землей, - многогранный объект для изучения социальности, роли хромосомных перестроек в видообразовании, и, что, возможно, самое интересное, - генетики детерминации пола. Для формирования гонад определяющим является время, место и уровень экспрессии ряда генов, список которых постепенно увеличивается, но ключевыми у плацентарных млекопитающих признаны Sry (Sex-determining Region Y) и SOX9 (SRY-box 9). Пять ныне живущих видов слепушонок рода Ellobius имеют три типа половых хромосом: XX-XY, X0-X0, XX-XX, при этом у видов, утративших Y-хромосому, нет гена Sry, и, как выяснилось сравнительно недавно [Bagheri-Fam, S., Sreenivasan, R., Bernard, P., Knower, K.C., Sekido, R., Lovell-Badge, R., Just, W. and Harley, V.R., 2012. Sox9 gene regulation and the loss of the XY/XX sex-determining mechanism in the mole vole Ellobius lutescens. Chromosome research, 20(1), pp. 191-199 http://link.springer.com/article/10.1007/s10577-011-9269-5], структура энхансера гена SOX9 нарушена из-за делеции в консервативном районе, что также должно вести к нарушению экспрессии. До сих пор непонятно, благодаря каким генам каскада детерминации пола выживают (и процветают!) обыкновенная и восточная слепушонки - фоновые широкоареальные виды слепушонки.
Но вернемся к половым хромосомам.
Изучение поведения двух X хромосом в мужском мейозе - нетривиальная задача. У человека подобные нарушения, как правило, проявляются в виде различных патологий (например, синдром де ля Шапеля), и могут быть сопряжены с транслокацией фрагмента Y хромосомы, несущего ген SRY, на X хромосому. Ранее мы показали [Kolomiets, O.L., Matveevsky, S.N. and Bakloushinskaya, I.Y., 2010. Sexual dimorphism in prophase I of meiosis in the Northern mole vole (Ellobius talpinus Pallas, 1770) with isomorphic (XX) chromosomes in males and females. Comparative Cytogenetics, 4, pp. 55-66. http://compcytogen.pensoft.net/articles.php?id=1693], что XX биваленты ведут себя по-разному у самцов и самок обыкновенной слепушонки: у самок в ооцитах на стадии лептотены хромосомы образуют особую конфигурацию, известную как «букет», тогда как в сперматоцитах «букет» не формируется; в ооцитах половые ХХ хромосомы синаптируют по всей длине полового бивалента, а у самцов неразличимые морфологически половые хромосомы "не считают себя" гомологичными и демонстрируют синапсис лишь в теломерных областях.
Анализу поведения половых XX хромосом в мейозе у самцов двух видов-двойников слепушонок, обыкновенной и восточной, на основе данных, полученных с помощью электронно-микроскопического и иммунофлуоресцентного исследования распластанных ядер сперматоцитов, и посвящена статья, вышедшая в Scientific Reports. Благодаря использованию новых методических подходов достигнута возможность изучения распределения от 5 до 8 антигенов в одном и том же ядре. Очевидно, что получение такой насыщенной "карты" активности белков, вовлеченных в процесс взаимодействия хромосом в мейозе особенно интересно для необычных половых хромосом. Что же удалось узнать? Установлено, что ХХ хромосомы самцов проявляют признаки функциональных различий, демонстрируя типично мужской тип поведения полового бивалента они формируют половое тельце. Между ХХ хромосомами образуются два коротких прителомерных участка синапсиса, в которых осуществляется рекомбинация, использование маркера локализации сайтов рекомбинации MLH1, одного из белков мисматч репарации, позволило показать это (см. рис. 2).
В центральной зоне полового бивалента на протяжении профазы I мейоза сохраняется зона асинапсиса между ХХ хромосомами, хроматин в которой подвергается транскрипционному сайленсингу.
Ранее нами было описано загадочное электронно-плотное аргирофильное ядрышкоподобное тельце, связанное с половым бивалентом. В новой работе впервые получены доказательства хроматиновой природы этого ядрышкоподобного тельца. Первым доказательством хроматиновой природы данной структуры является интенсивное окрашивание её красителем Dapi. Вторым - связывание с ней белков сайленсинга хроматина. Также было показано, что распределение ряда маркеров инактивации хроматина полового тельца отличается у слепушонок от других млекопитающих.
Проявление типичных для XY полового бивалента особенностей поведения в мейозе в случае XX полового бивалента у самцов слепушонок двух видов может быть первым шагом в эволюции половых хромосом от морфологически идентичных к гетероморфным.
У слепушонок с XX хромосомами у обоих полов морфологическая идентичность маскирует функциональный гетероморфизм половых хромосом, который удается увидеть только в мейозе. Возможно, что после потери Y хромосомы произошла дупликация оставшейся X хромосомы, и в настоящее время мы наблюдаем первый этап формирования гетероморфных половых хромосом.
Существование таких групп как слепушонки, заставляет усомниться в единственности возможного пути детерминации пола у млекопитающих и заново оценить роль общепринятых ключевых факторов детерминации пола.

Рис. 1. Восточная слепушонка Ellobius tancrei
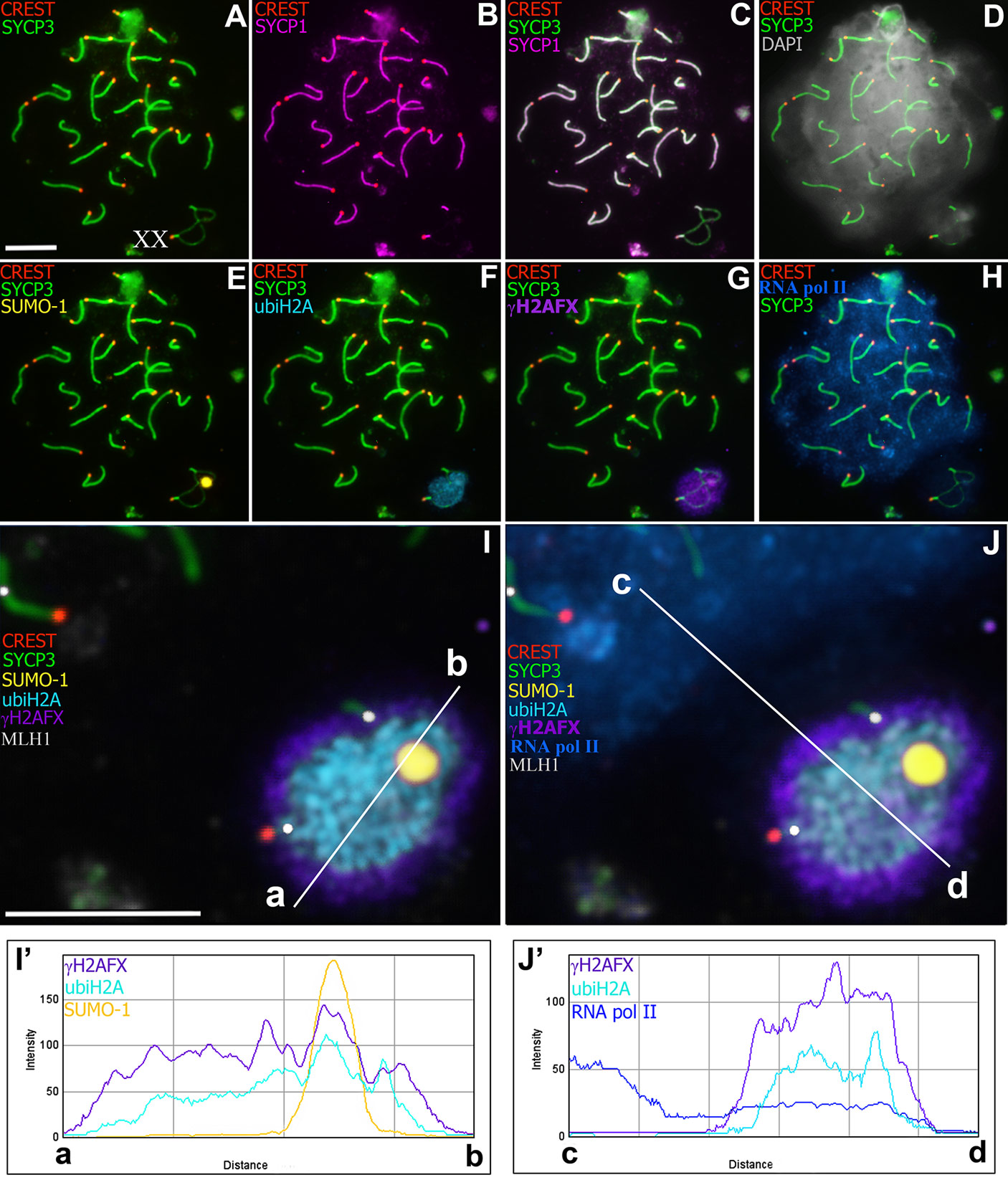
Рис. 2. Сперматоциты Ellobius talpinus, пахитена. Локализация SCP3 (green), SCP1
(magenta), CREST (red), SUMO-1 (yellow), ubiH2A (cyan), Polo II (blue), γH2AX (violet),
MLH1 (gray) and DAPI configuration (gray). A-H. Один и тот же сперматоцит иммуноокрашен
разными антителами. I-J. Половой (ХХ) бивалент (from figs A-H). В зоне теломерного синапсиса
наблюдаются MLH1-сигналы, в зоне асинапсиса локализованы белки инактивации хроматина. SUMO-1
расположен в пределах ядрышко-подобного тельца (ЯПТ). Polo II интенсивно светится в основной
части сперматоцита, а в пределах полового бивалента дает слабый сигнал. Взаиморасположение
SUMO-1, ubiH2A, Polo II, γH2AX отражено на графиках a-b и c-d (see I’
and J’).
(рис. 3 из обсуждаемой статьи)
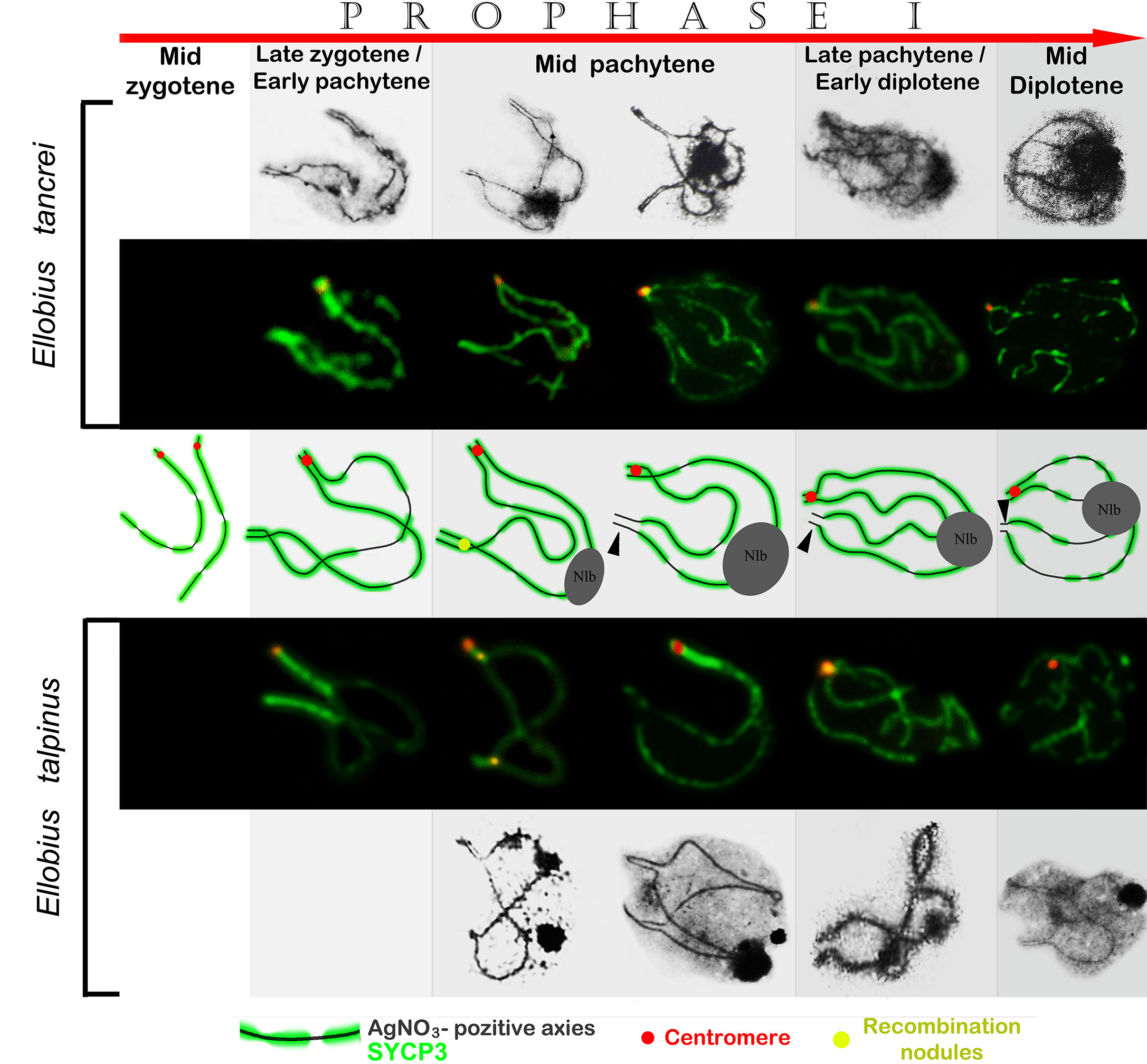
Рис. 3. Половой XX бивалент в профазе мейоза у двух видов слепушонок,
восточной Ellobius tancrei и обыкновенной, E. talpinus (рис. S2 из дополнительных
материалов к статье)
Copyright © 2016 Bakloushinskaya I.Yu.