 © Куликова Дина Александровна, с.н.с., к.б.н. лаборатории МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИХ ПРОЦЕССОВ РАЗВИТИЯ
© Куликова Дина Александровна, с.н.с., к.б.н. лаборатории МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИХ ПРОЦЕССОВ РАЗВИТИЯМобильные элементы – это подвижные участки генома. Их транспозиции могут менять структуру генов и их экспрессию. МЭ занимают, в среднем, около 50% генома эукариот и являются одной из ведущих движущих сил эволюции при условии их перемещения de novo в клетках зародышевого пути. МЭ могут перемещаться и в соматических тканях, в том числе в нервной. Однако, изучать индивидуальные транспозиции с помощью полногеномного секвенирования невозможно, поскольку при этом отсутствует учёт индивидуальных различий между клетками. В связи с этим в исследованиях принято говорить о корреляции между нейродегенерацией и изменением уровня экспрессии транспозона. Однако, не всегда повышение уровня экспрессии транспозона связано с его мобилизацией и перемещением. Для понимания механизмов изменений в работе индивидуальных нервных клеток необходимо выяснить, что контролирует экспрессию последовательностей, соответствующих МЭ. Часто МЭ сидят в интронах генов, привнося туда сайты сплайсинга, и в результате образуется химерный транскрипт, состоящий из мРНК транспозона и мРНК соответствующего гена. Около 4% генов человека несут в себе последовательности транспозонов в виде новых экзонов, а 75% длинной некодирующией РНК человека содержат их фрагменты. Однако, до сих пор неясно какой вклад такие химерные мРНК вносят в общий пул мРНК транспозонов в соматических клетках. Чтобы оценить распространенность химерных транскриптов в геноме необходимо понять, связана ли экспрессия каждого транспозона в геноме с экспрессией окружающих его генов. Недавно появившаяся технология высокопроизводительного секвенирования транкриптома отдельных клеток делают возможным такую оценку. Авторы поставили задачу оценить какую долю составляют химерные транскрипты в группах клеток среднего мозга мух. Для этого они создали новые инструменты анализа и получили необычные результаты.
Обычно при секвенировании количественная оценка экспрессии транспозона происходит при нормировании на количество его копий на геном, но эталонный геном значительно отличается по количеству вставок от генома отдельных линий, что делает количественную оценку неточной. Авторы, применяя новую методику анализа данных scRNA-seq из отдельных клеток среднего мозга мух, картировали транспозоны, находящиеся внутри генов и в межгенном пространстве и показали, что экспрессия транспозонов, находящихся внутри определенных генов, коррелирует с экспрессией этих генов. Кроме того, их экспрессия выявлялась в определенных клетках мозга (рис. 1).
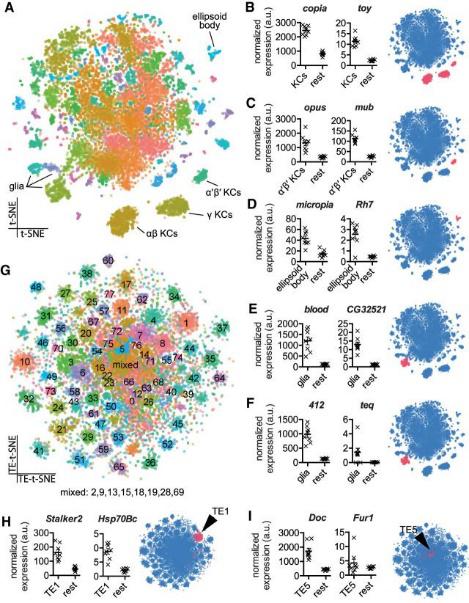
C помощью разработанного авторами метода, основанного на принципе Хайди-Вайнберга, стало возможно количественно оценить влияние экспрессии гена на экспрессию транспозона, который в нем находится. Для всех транспозонов была обнаружена хотя бы одна копия, которая находилась внутри гена и показывала коррелированный паттерн экспрессии (рис. 2). Исключениями являются теломерные мобильные элементы TART, TART-A и HeT-A, которые, скорее всего, всегда экспрессируются автономно.
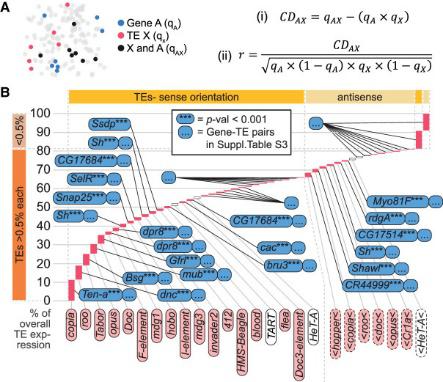
В статье показано, что мРНК транспозонов образуют множественные химеры с пре-мРНК генов и отличить эти транскрипты от собственных транскриптов транспозонов обычными методами невозможно.
Кроме того, авторы разработали метод учета артефактных химерных транскриптов, которые могут случайно формироваться при амплификации ДНК in vitro и предложили использовать коэффициент ложного обнаружения.
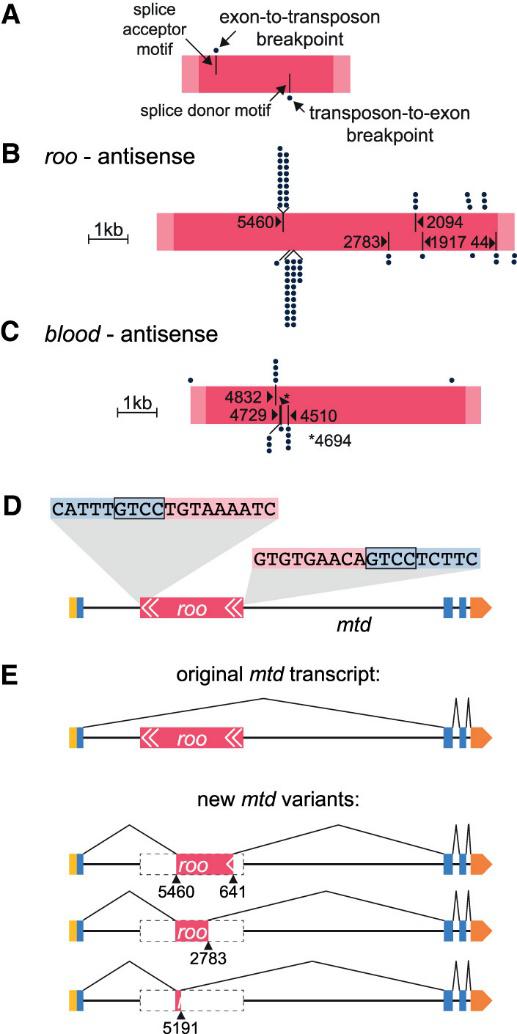
Исследуя химерные транскрипты, авторы обнаружили, что транспозоны могут вносить в мРНК хозяина альтернативные сайты сплайсинга (Рис. 3). Причем почти для всех подсемейств транспозонов был обнаружен хотя бы один нейрональный ген, в транскриптах которого есть химеры ген-транспозон. Иногда эти новые сплайс-сайты могут способствовать удалению одного из хозяйских экзонов. Интересно, что один транспозон может вносить несколько альтернативных сплайс-сайтов. Эти новые сплайс-сайты распознаются слайсосомами, что приводит к разнообразию изоформ нейрональных генов.
Проанализировав количественный вклад химерных транскриптов в общий пул транскриптов, авторы показали, что, по крайней мере для некоторых генов большинство транскриптов содержали последовательности транспозонов. Так 95,3% трансткриптов гена Rhodopsin 7 имеют вставку LTR ретротранспозона micropia в 3' UTR, а все мРНК гена Allatostatin C receptor 1 (AstC-R1) содержали участок F-элемента, который мог сплайсироваться в один из двух разных акцепторных сайтов сплайсинга гена. Авторы описывают еще ряд генов, транскрипты которых содержат вставки целых мобильных элементов или их части.
Сравнение пулов химерных транскриптов из разных линий дрозофилы показало, что большая часть из них уникальна для каждой линии, а некоторые транскрипты уникальны даже для индивидуальных особей.
На основании своего анализы авторы приходят к выводу, что транспозоны в нервной ткани вносят значительный вклад в разнообразие транскриптов кодирующих белок генов и расширяют репертуар форм белков, которые могут менять работу групп клеток в пределах одного организма. Эти изменения не передаются по наследству, поскольку происходят в соматической ткани. Тем не менее, они могут стать причиной девиантных форм поведения и различных нейропатологий у отдельных индивидов.
Новость подготовила © Куликова Д.А.
25.03.2021