 © Сорокина Светлана Юрьевна,
к.б.н., с.н.с. лаб. Эволюционной генетики развития ИБР РАН, Область научных интересов: генетическая структура вида, изменчивость митохондриальной ДНК, группа близкородственных видов дрозофил virilis
© Сорокина Светлана Юрьевна,
к.б.н., с.н.с. лаб. Эволюционной генетики развития ИБР РАН, Область научных интересов: генетическая структура вида, изменчивость митохондриальной ДНК, группа близкородственных видов дрозофил virilis
Филогенетические отношения рода Homo представляют для нас особый интерес не только потому, что мы относимся к нему как вид, но еще и потому, что наш вид – последний в этом роде. Информацию о наших предках, близких и далеких родственниках приходится собирать по разрозненным палеонтологическим останкам давно вымерших видов. Еще совсем недавно, считалось, что у H. sapiens был один сестринский вид, живший с ним в одно время и состоявший с ним в конкурентных отношениях в период расселения по Евразии – H. neanderthalensis. Филогенетические отношения не вызывали вопросов: две ветви рода Homo - одна в Европе, другая в Африке, с хорошо выраженными отличиями в морфологии и несколько различающимися временами дивергенции по ядерным и митохондриальным геномам, что можно было объяснить разницей в скоростях эволюции данных маркеров. Но как только появилась информация еще об одном нашем родственнике – H. denisova, схема усложнилась гораздо сильнее, чем можно было ожидать. В системе из трех элементов не только время дивергенции, но и порядок ветвления ядерных и митохондриальных деревьев оказался различным, указывая на то, что истории таксонов переплетались если не тесно, то достаточно, для того, чтобы оставить отпечатки событий интрогрессии в генофондах современных и древних людей. По мере изучения полных геномов постепенно открываются все новые факты неродственных гибридизаций, в которые оказываются вовлечены не только известные, но и пока еще неизвестные более или менее удаленные представители рода Homo. Показано, что некоторые коренные африканские народы имеют в своем геноме признаки гибридизации с неизвестной разновидностью, отделившейся от сапиенсной ветви вскоре после разделения сапиенсной и неандертало-денисовской ветвей (степень родства между H. sapiens и этой разновидностью такая же, либо чуть меньше, чем между H. neanderthalensis и H. denisova, и таким образом это самый близкий наш родственник на сегодняшний день) [1]. В геноме денисовца есть свидетельства гибридизации с удаленной разновидностью, дивергировавшей от основного ствола Homo около 1 млн. лет назад [2]. С этим фактом согласуется необычно удаленный от H. sapiens и H. neanderthalensis митохондриальный гаплотип денисовского человека, предположительно полученный им в результате интрогрессии [3]. Однако данная гипотеза имеет ряд недостатков, которые пока не представляется возможным объяснить. Например, почему в древних останках из пещеры Sima de los Huesos, принадлежавших по данным ядерной ДНК разновидности, от которой произошли неандертальцы, обнаруживается мтДНК денисовской ветви? Тоже интрогрессия? Вероятность двух независимых случайных событий значительно ниже, чем одного... И хотя этот вопрос по-прежнему остается загадкой, даже принимая во внимание только геномные данные, становится понятно, что у наших предков были еще современники, кроме человека неандертальского и человека денисовского.
В начале мая в журнале Nature вышла публикация международного коллектива авторов, которые применили подход, основанный на масс-спектрометрическом анализе палеопротеома из костных останков [4], [5], на биологическом материале из фрагмента нижней челюсти, найденного в пещере Байшия (Тибетское нагорье, провинция Сяхе, Китай) на высоте 3280 м еще в 1980 г. (Рис. 1). Образец по морфологическим признакам не мог быть отнесен к H. sapiens или H. neanderthalensis и принадлежал особи, жившей в период среднего плейстоцена 160 тыс. лет назад в тех же местах, где в настоящее время проживает коренное население Тибета. Исследуемый фрагмент нижней челюсти (образец Xiahe) имеет ряд общих черт (в частности отсутствие третьего моляра) с аналогичными находками на территории Восточной Азии, также относящимися к среднему плейстоцену. Как и ожидалось, из-за возраста и особенностей климата ДНК в образце не сохранилась. Однако авторам удалось извлечь белки из дентина (основной ткани зуба) одного из сохранившихся моляров. Несмотря на сильную деградацию пептидных цепей, авторы собрали и определили аминокислотные последовательности протяженных участков двух коллагенов COL1α1 и COL1α2 и еще четыре коллагена (COL2α1, COL3α1, COL5α1 и COL5α2) удалось собрать только фрагментарно. В след за этим было проведено сравнение полученных данных с соответствующими участками геномов известных гоминид, транслированными в аминокислотые последовательности, в результате чего удалось установить принадлежность древних останков представителю денисовской ветви (Рис. 2). Данное открытие позволяет сделать сразу несколько важных выводов:
1) Денисовцы жили на больших высотах (выше 3 тыс. м) и были приспособлены к условиям гипоксии задолго до современных жителей Тибета. Это объясняет выявленную ранее интрогрессию денисовской формы гена EPAS1 - транскрипционного фактора, ответственного за регуляцию функций кровеносной системы в условиях снижения уровня кислорода, у тибетцев [6]. До этого адаптивная значимость данного аллеля для денисовцев была неочевидна, так как Денисовская пещера, где были обнаружены все известные до сих пор останки этой разновидности Homo, находится на высоте всего лишь 700 метров, и особых адаптаций к условиям высокогорья ее обитателям не требовалось.
2) Учитывая то, что до сих пор нам практически ничего не было известно о морфологии Денисовца, новые данные о строении его нижней челюсти можно считать существенным достижением. Полученные сведения позволяют предполагать принадлежность некоторых морфологически сходных образцов из Восточной Азии к этой разновидности рода Homo, что, в свою очередь, позволит делать выводы о филогеографической истории таксона.
3) Подход, представленный в данной статье, открывает широкие возможности для исследования палеоматериала и позволяет определять филогенетическое родство в случаях, когда выделить ДНК из образца не удается, т.е. по более древним останкам, а также по останкам из районов с неподходящим для сохранения ДНК климатом. Например, было бы очень интересно узнать отношение к нашему виду африканской разновидности H. naledi [7], загадочного “хоббита” с острова Флорес (H. floresiensis) [8], а также недавно обнаруженного на Филиппинах нового представителя рода Homo – H. luzonensis [9].
Таким образом, коллагены дентина в ближайшем будущем могут оказаться той баркодинговой последовательностью, которая позволит систематизировать спектр разнообразных гоминид, известных по хорошо сохранившимся, но утратившим ДНК останкам, дополнить его новыми представителями по данным отдельных фрагментов скелетов, таксономическую идентичность которых трудно или невозможно установить морфологически, и объединить все это с уже известными геномными данными. В результате мы получим наиболее полную молекулярную генеалогию нашего рода и сможем, наконец, узнать, “кто все эти люди” (Род Homo – люди).
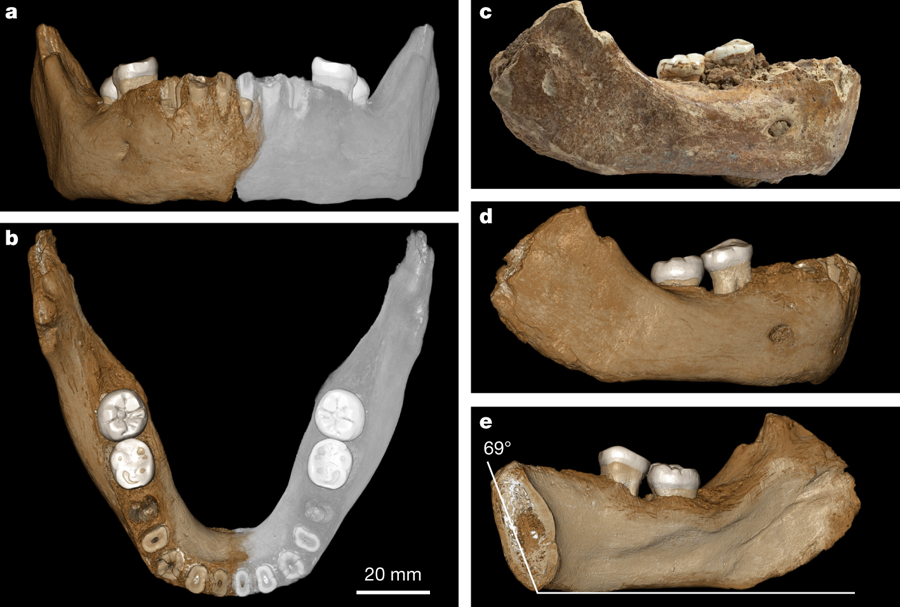
Рис.1 Правая половина нижней челюсти с двумя сохранившимися молярами, найденная в пещере Байшия (Baishiya Karst Cave) в провинции Сяхе (Xiahe) на северо-востоке Тибетского нагорья в 1980г. Возраст по данным радиоизотопного анализа (метод урановых серий) – 160 тыс. лет.
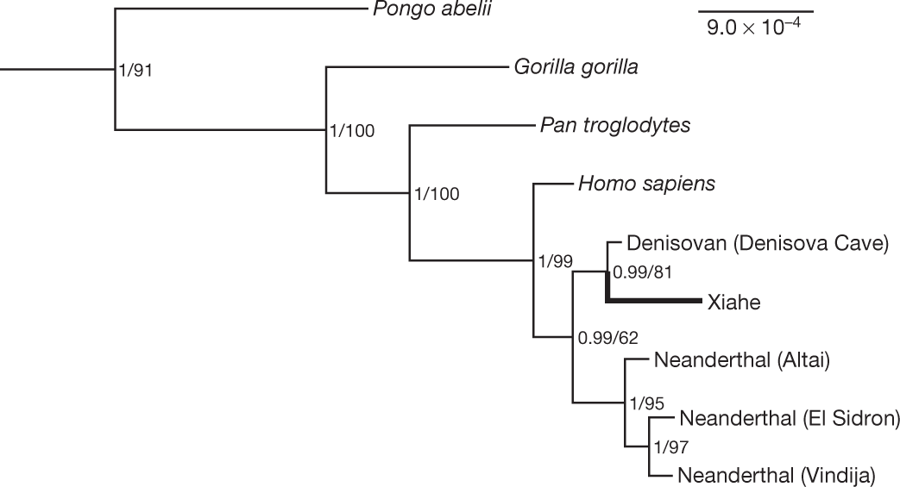
Рис. 2. Положение образца из пещеры Xiahe на древе гоминид по данным аминокислотных последовательностей шести коллагенов дентина (COL1α1, COL1α2, COL2α1, COL3α1, COL5α1 и COL5α2).
1) Lorente-Galdos B., Lao O., Serra-Vidal G., Santpere G., Kuderna L.F.K., Arauna L.R., Fadhlaoui-Zid K., Pimenoff V.N., Soodyall H., Zalloua P., Marques-Bonet T., Comas D. Whole-genome sequence analysis of a Pan African set of samples reveals archaic gene flow from an extinct basal population of modern humans into sub-Saharan populations// Genome Biol. 2019 Apr 26;20(1):77. doi: 10.1186/s13059-019-1684-5.
2) Pääbo S. The diverse origins of the human gene pool// Nat Rev Genet. 2015 Jun;16(6):313-4
3) Krause J, Fu Q, Good JM, Viola B, Shunkov MV, Derevianko AP, Pääbo S. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia// Nature. 2010 Apr 8;464(7290):894-7. doi: 10.1038/nature08976. Epub 2010 Mar 24.
4) Welker, F. et al. Palaeoproteomic evidence identifes archaic hominins associated with the Châtelperronian at the Grotte du Renne// Proc. Natl Acad. Sci. USA 113, 11162–11167 (2016).
5) Welker, F., Soressi, M., Rendu, W., Hublin, J.-J. & Collins, M. J. Using ZooMS to identify fragmentary bone from the late Middle/Early Upper Palaeolithic sequence of Les Cottés, France// J. Archaeol. Sci. 54, 279–286 (2015)
6) Huerta-Sánchez, E. et al. Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA// Nature 512, 194–197 (2014)
7) Berger LR, et al. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa// Elife. 2015 Sep 10;4. doi: 10.7554/eLife.09560.
8) Brown P, Sutikna T, Morwood MJ, Soejono RP, Jatmiko, Saptomo EW, Due RA.A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia// Nature. 2004 Oct 28;431(7012):1055-61.
9) Detroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, Robles E, Grün R, Piper PJ. A new species of Homo from the Late Pleistocene of the Philippines. Nature. 2019 Apr;568(7751):181-186. doi: 10.1038/s41586-019-1067-9. Epub 2019 Apr 10.
Новость подготовила © Сорокина С.Ю.
23.05.2019