Dorkenwald, S., Matsliah, A., Sterling, A.R. et al. Neuronal wiring diagram of an adult brain. Nature 634, 124–138 (2024). https://doi.org/10.1038/s41586-024-07558-y
Eckstein N, Bates AS, Champion A, et al., Neurotransmitter classification from electron microscopy images at synaptic sites in Drosophila melanogaster. Cell. 2024 May 9;187(10):2574-2594.e23.doi: 10.1016/j.cell.2024.03.016
Lin, A., Yang, R., Dorkenwald, S. et al. Network statistics of the whole-brain connectome of Drosophila. Nature 634, 153–165 (2024). https://doi.org/10.1038/s41586-024-07968-y
Schlegel, P., Yin, Y., Bates, A.S. et al. Whole-brain annotation and multi-connectome cell typing of Drosophila. Nature 634, 139–152 (2024).https://doi.org/10.1038/s41586-024-07686-5

Область научных интересов: нейроэтология, нейробиология, нейротрансмиттеры, поведение.
11 ноября ушел из жизни Дмитрий Антонович Сахаров (1930-2024), уникальный человек, наставник, ученый и поэт. В этом же году мировое сообщество отмечает 50-летие выхода в свет его книги «Генеалогия нейронов», оказавшей громадное влияние на несколько поколений нейробиологов. В 2024 году вышли статьи по нейрогенезу и созданию траснкриптомного атласа мозга, позволяющие проверить некоторые гипотезы Д.А. Сахарова, прежде всего его идею о связи нейротрансмиттерных фенотипов с онто- и филогенетическими линиями развития нейронов.

По-видимому, главный вопрос, интересовавший Д.А. Сахарова, это причина разнообразия нейротрансмиттеров в нервных системах, который он рассматривал в эволюционном, онтогенетическиом и функциональном аспекте. Д.А. Сахаров является автором оригинальной гипотезы о полигенезе нейронов, или множественном происхождении нейронов в эволюции из разных тканевых и клеточных предшественников. Гипотеза впервые была сформулирована в 1970 г. (Сахаров 1970) и подробно рассматривается в книге «Генеалогия нейронов» 1974, которой в этом году исполнилось 50 лет. Множественность трансмиттерных фенотипов во всех изученных к тому времени нервных системах он объяснял в первую очередь разными линиями происхождения разных нейронов или полигенезом. За прошедшие годы появилось много геномных и транскриптомных исследований, которые подтверждают и то, что нейрон мог не один раз возникать в эволюции (наиболее яркий пример здесь нервные системы грибневиков), и то, что все исследованные на сегодняшний день нервные системы представлены нейронами разной трансмиттерной специфичности (Moroz 2021; Moroz et al., 2021b). Однако впоследствии его интерес явно сместился в сторону функциональной роли такой множественности нейротрансмиттеров. Он пришел сначала к интуитивному пониманию того, что в нервной системе огромную роль должна играть несинаптическая коммуникаций между нейронами, при которой упорядоченность взаимодействий достигается за счет наличия соответствующих рецепторов у соответствующих нейронов. Это представление было озвучено в 1985-1990 гг под названием «гетерон» (Сахаров 1985; 1990). Уже к девяностым годам накопилось много экспериментальных доказательств объемной нейротрансмиссии, и появились другие авторы, активно развивающие это направление (Fuxe et al., 1990; Benfenati, Agnati 1991; Bach-y-Rita P, Illis 1993). Однако отличие взглядов Д.А. Сахарова от их представлений о роли несинаптической нейротрансмиссии было принципиальным. Он рассматривал объемную нейрональную секрецию не как внешнюю модуляцию жесткого синаптического ансамбля, а как основу упорядоченного взаимодействия нейронов на всех уровнях организации нервной системы. А классический синапс с его изолирующими барьерами рассматривал как редкий и предельный случай такой коммуникации.
Стремительное развитие транскриптомики одиночных клеток, позволяющей не только устанавливать онтогенетическое родство нейронов на основании сходства их транскриптомов, но и прослеживать развитие разных линий в онтогенезе, может ответить на вопрос о о связи нейротрансмиттерных фенотипов с линиями развития нейронов. Некоторые важные для гипотезы полигенеза данные уже получены в 2023-2024 годах при построении транскриптомных клеточных атласов мозга мыши и дрозофилы.
Наиболее значимые подтверждения гипотезы Д. Сахарова о связи происхождения нейрона и его трансмиттерного фенотипа получены на дрозофиле. Первая работа, выполненная на брюшной нервной цепочке, вышла в 2019 с четким выводом, обозначенном в самом названии статьи «Neurotransmitter identity is acquired in a lineage-restricted manner in the Drosophila CNS» (Lacin et al., 2019). Авторы создали полную карту трех нейротрансмиттерных типов (ацетилхолин, ГАМК или глутамат) для всей брюшной нервной цепочки в соответствии с их происхождением, т.е. отношением к одной из 32 известных гемилиний развития. Они не обнаружили ни одного случая использования нейронами более одного нейротрансмиттера, хотя ацетилхолин-специфический ген ChAT транскрибируется во многих глутаматергических и ГАМКергических нейронах, но эти транскрипты обычно не покидают ядро и не транслируются. Наиболее важным результатом своей работы авторы сочли формулировку простого правила: все нейроны в пределах онтогенетической гемилинии используют один и тот же нейротрансмиттер. Таким образом, идентичность нейротрансмиттера приобретается на уровне стволовых клеток. Это правило немедленно приобрело название «Правило Лацина» (Lacin’s law). Не стоит, наверное, ожидать от международного сообщества памяти о вышедших, в том числе в международных журналах (Sakharov 1974a,b) работах 50 летней давности Д.А. Сахарова, выдвинувшем гипотезу о связи трансмиттерного фенотипа с происхождением нейрона.
Правило Лацина заинтересовало некоторых исследователей, правильность этого постулата проверили и по отношению к головному мозгу дрозофилы, а затем и по отношению к мозгу млекопитающих. Для мозга дрозофилы правило выполнялось, хоть с некоторыми оговорками. Так в мае 2024 году в журнале Cell вышла статья, в которой искусственные нейронные сети научили предсказывать трансмиттерный фенотип нейронов для шести нейротрансмиттеров (ацетилхолин, глутамат, ГАМК, серотонин, дофамин, октопамин) по электронно-микроскопическим фотографиям секретирующих окончаний. Точность такого предсказания достигала 87% для отдельных синапсов, 94% для нейронов и 91% для известных типов клеток во всем мозге D. melanogaster. Способность нейронных сетей делать такие предсказания свидетельствует о том, что существуют тонкие, но существенные различия между фенотипами трансмиттеров по морфологии секретирующих окончаний нейрона. Анализируя распределение нейротрансмиттеров в мозге, авторы показали, что нейроны, которые развиваются вместе, в основном экспрессируют только один быстродействующий трансмиттер (ацетилхолин, глутамат или ГАМК), то есть подтвердили правило Лацина.
Октябрь 2024 отмечен мощным выступлением консорциума по изучению мозга дрозофилы, выпустившим сразу три статьи в Nature (Dorkenwald et al., 2024; Lin et al., 2024; Schlegel et al 2024). Они посвящены построению транскриптомного, коннектомного и онтогенетического атласа мозга дрозофилы. Эти данные позволили максимально уточнить выявленные выше закономерности формирования трансмиттерного фенотипа нейронов. Показано, что примерно 120 идентифицированных нейробласта генерируют все нейроны мозга и часть зрительных проекционных нейронов в каждом (левом и правом) полушарии. Каждая из этих стволовых клеток определяется уникальным транскрипционным кодом и генерирует стереотипную линию в точном порядке путем асимметричного деления. Каждый нейробласт обычно производит две гемилинии, которые заметно отличаются по нейронной морфологии и могут экспрессировать разные нейротрансмиттеры, но нейроны в каждой гемилинии обычно экспрессируют один быстродействующий трансмиттер. Внутри гемилинии нейроны образуют отростки, которые собираются вместе в один плотный пучок, образуя общий тракт, который входит, пересекает и соединяет разные области нейропиля. Таким образом, не нейробласт, а именно гемилинии представляют собой естественную функциональную, а также эволюционную группировку, с помощью которой можно изучать нервную систему. В этом выводе авторов представляется очень важным и близким гипотезе Д.А. Сахарова акцент не только на онтогенетической систематике нейронов, объединенным общим химизмом, но и на эволюционной составляющей, которая предполагает наследуемость трансмиттерных фенотипов.
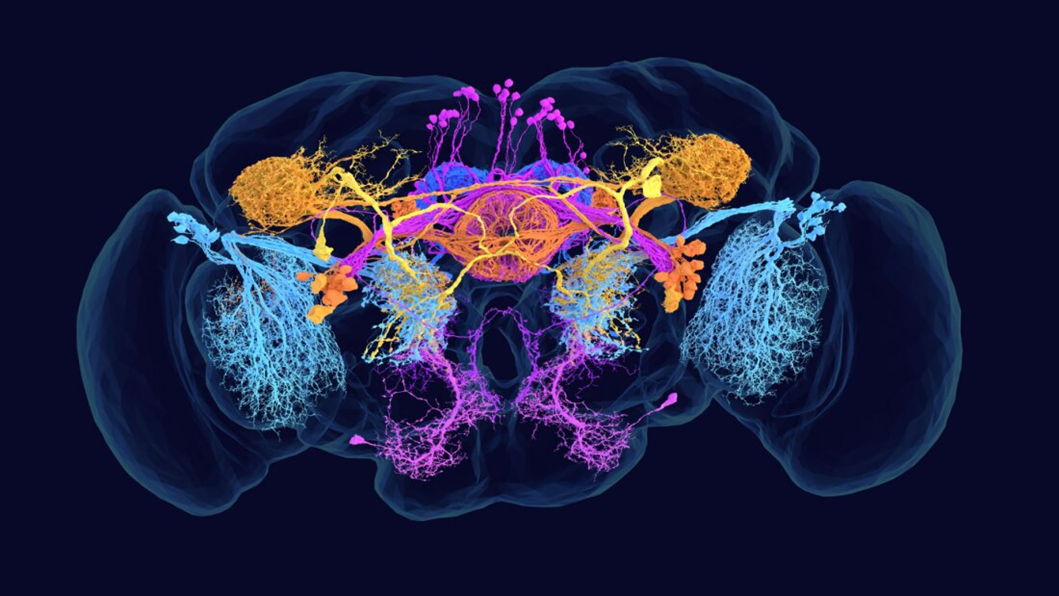
Amy Sterling for FlyWire, Princeton University, (Dorkenwald et al., Nature, 2024), нейроны мозга дрозофилы, шести разных трансмиттерных типов.
В отличие от дрозофилы, анализ связи трансмиттерного фенотипа с происхождением нейрона у млекопитающих выявил довольно смешанную картину (см. комментарий Дьяконова 2024 «На основе пространственной транскриптомики с разрешением в одну клетку создан атлас всего мозга мыши»). С одной стороны, есть сигнальные молекулы (и классические нейротрансмиттеры, и пептиды), которые экспрессируются довольно строго в соответствии с линиями развития нейронов. Но на этом фоне глутамат, ГАМК и некоторые нейропептиды экспрессируются настолько широко и разнообразно, что правило «общее происхождение – общий нейротрансмиттер» кажется явно нарушенным.
Такая картина предполагает многофакторность формирования трансмиттерного фенотипа нейронов у млекопитающих. То есть, помимо происхождения нейрона в эволюции и онтогенезе, на формирование трансмиттер-специфических нейронов оказывали сильное влияние и какие-то другие факторы. Одним из важных факторов может быть уже упомянутая функциональность гетерохимизма: функциональный ансамбль не может работать на одинаковых нейронах, если он весь состоит из нейронов общего происхождения. (Под ансамблем понимается саморегулирующаяся система нейронов с обратными связями, способная контролировать определенную функцию, хороший пример разнообразные центральные генераторы паттерна). Между тем, в связи с возросшей нагрузкой на стабильность нейронального генома у теплокровных (Дьяконова 2023), в их эволюции могла возникнуть необходимость быстро увеличить число нейронов за счет увеличения одной популяции нейронов общего происхождения. В таком случае формирование гетерохимизма такой популяции могло происходить уже вторично. Интересно, что «минимальный гетерохимизм» формируется, главным образом, за счет глутамат- и ГАМКергических нейронов. Работы 2021 года позволяют предположить, почему (Moroz 2021; Moroz et al., 2021). Благодаря высокому содержанию глутамата как метаболита во всех живых клетках, все, что требуется для формирования его трансмиттерной функции, это разблокировка экспрессии глутаматного везикулярного транспортера. Сходная «эпигенетическая простота» просматривается и в отношении ГАМК, которая синтезируется из глутамата. Здесь достаточно всего двух генов для формирования трансмиттерной функции.
Кроме того, на формирование трансмиттерного ландшафта мозга в эволюции могла влиять разная биологическая стоимость разных трансмиттерных фенотипов и в терминах энергии (Moroz et al., 2021), и в терминах опасности для стабильности генома нейрона (Дьяконова 2022; 2023; Dyakonova 2020). Действительно, с энергетической точки зрения глутамат является «дешевым» трансмиттером. Он требует минимальных затрат на синтез и позволяет к тому же использовать его метаболиты в качестве источника энергии (Moroz et al., 2021). Эпигенетической простотой и энергетической дешевизной глутамата как нейротрансмиттера можно объяснять выраженное увеличение доли глутаматергических нейронов головного мозга в эволюции млекопитающих (от 50 процентов у мыши до 80 у человека, в сравнении с первичноротыми, у которых не более 10 процентов нейронов являются глутаматергическими) (Moroz et al., 2021). Можно предположить, что мозг стремительно увеличивался в эволюции млекопитающих, главным образом, за счет увеличения доли глутаматергических нейронов и, что вторичное обогащение трансмиттерного разнообразия новых популяций нейронов происходило за счет ГАМК и некоторых пептидов.
Таким образом, разные факторы в совокупности формировали эволюцию мозга, их сложное взаимодействие может объяснять довольно неоднозначную связь происхождения нейрона и его химического фенотипа у млекопитающих, в отличие от некоторых первичноротых животных.
В заключение следует сказать, что эта тема еще не закрыта, напротив, мы являемся свидетелями ее начавшегося возрождения на новом этапе развития научных технологий. В ближайшие годы нас ждут данные по закономерностям формирования нейронального разнообразия в разных и отдаленных эволюционных группах животных, которые, существенно прояснят справедливость гипотез Д.А. Сахарова.
Литература:
- Дьяконова В.Е. (2022). Происхождение и эволюция нервной системы: новые данные сравнительных полногеномных исследований многоклеточных животных, Онтогенез, 53 (1), 63-74.
- Дьяконова В.Е. Нестабильная ДНК нейрона: счетчик продолжительности жизни и драйвер эволюции. Биохимия. (2023) № 11.DOI: 10.1134/S0006297923110044
- Сахаров, Д. А. (1970). Основания к построению системы нервных клеток. Журнал общей биологии, 31 (4), 449–457.
- Сахаров, Д. А. (1974). Генеалогия нейронов. М.: Наука.
- Сахаров, Д. А. (1985). Синаптическая и бессинаптическая модели нейронной системы. Простые нервные системы. Ч. 2. Казань: КГУ, 78–80.
- Сахаров, Д. А. (1990). Множественность нейротрансмиттеров: функциональное значение. Журнал эвoлюционной биoxимии и физиoлогии, 26 (5), 733–741.
- Chen N, Zhang Y, Rivera-Rodriguez EJ, Yu AD, Hobin M, Rosbash M, Griffith LC. Widespread posttranscriptional regulation of cotransmission. Sci Adv. 2023 Jun 2;9(22):eadg9836. doi: 10.1126/sciadv.adg9836.
- Dorkenwald, S., Matsliah, A., Sterling, A.R. et al. Neuronal wiring diagram of an adult brain. Nature 634, 124–138 (2024). https://doi.org/10.1038/s41586-024-07558-y
- Lacin H., Chen H.-M., Long X., Singer R.H., Lee T., Truman J.W. Neurotransmitter identity is acquired in a lineage-restricted manner in the Drosophila CNS. Elife. 2019;8 doi: 10.7554/eLife.43701.
- Lin, A., Yang, R., Dorkenwald, S. et al. Network statistics of the whole-brain connectome of Drosophila. Nature 634, 153–165 (2024). https://doi.org/10.1038/s41586-024-07968-y
- Moroz, L. L. (2021). Multiple origins of neurons from secretory cells. Frontiers in Cell and Developmental Biology, 9, 669087.
- Moroz, L. L., Nikitin, M. A., Poličar, P. G., Kohn, A. B., Romanova, D. Y. (2021a). Evolution of glutamatergic signaling and synapses. Neuropharmacology, 199, 108740.
- Moroz, L. L., Romanova, D. Y., Kohn, A. B. (2021b). Neural versus alternative integrative systems: molecular insights into origins of neurotransmitters. Philosophical Transactions of the Royal Society B, 376 (1821), 20190762.
- Sakharov DA. Proceedings: Evolutionary aspects of transmitter heterogeneity. J Neural Transm. 1974а; Suppl 11(0):43-59. PMID: 4152422.
- Sakharov, D. A. (1974b). Evolutionary aspects of transmitter heterogeneity. In Neurovegetative Transmission Mechanisms: Proceedings of the International Neurovegetative Symposium, Tihany, June 19–24, 43–59. Vienna: Springer Vienna.
- Schlegel, P., Yin, Y., Bates, A.S. et al. Whole-brain annotation and multi-connectome cell typing of Drosophila. Nature 634, 139–152 (2024). https://doi.org/10.1038/s41586-024-07686-5
- Yao Z, van Velthoven CTJ, Kunst M, Zhang M, McMillen D, Lee C, et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature. 2023 Dec;624(7991):317-332. doi: 10.1038/s41586-023-06812-z.
Новость подготовила
© В.Е.Дьяконова
22.12.2024