 © Симонова Ольга Борисовна,
доктор биологических наук, руководитель лаборатории молекулярно-генетических процессов развития ИБР РАН.
© Симонова Ольга Борисовна,
доктор биологических наук, руководитель лаборатории молекулярно-генетических процессов развития ИБР РАН.
Новое исследование, выполненное под руководством ученых из Чикагского университета, ставит под сомнение одно из классических предположений о том, как появляются новые гены. Исследования показывают, что случайные, некодирующие (бессмысленные) участки ДНК могут быстро эволюционировать с образованием новых белок-кодирующих генов. Эти гены de novo, или гены «с нуля», становятся материалом для эволюционного отбора, способствуя образованию новых видов и биоразнообразию.
В течение десятилетий ученые считали, что существуют два основных пути возникновения новых генов: 1) дупликация исходного гена с последующей дивергенцией паралогов и 2) рекомбинация участков гена, кодирующих разные белковые домены. Если одна из копий гена затем дивергирует и меняет свою функциональность, то она становится новым геном. Отбор поддерживает такое событие, если исходный ген выполнял две разные функции, «работал на два фронта», поскольку дупликация способствует разделению функций между его копиями и повышению эффективности работы обоих генов. При рекомбинации фрагменты генетического материала (кодирующие разные белковые домены) перетасовываются для создания новых комбинаций и новых генов.
Ученые давно задаются вопросом о третьем механизме, когда гены могли бы возникать «с нуля» - de novo. Мы сейчас знаем, что у всех организмов есть протяжённые участки генома, которые не кодируют белки. Могут ли эти некодирующие участки приобретать мутации, которые внезапно делают их функциональными? Каков механизм возникновения нового гена «с нуля»: из уже существующих транскриптов (длинных некодирующих РНК) или исходным сырьем для них являются открытые рамками считывания (ORF), так называемые «прото-гены», которые случайно формируются в геноме? Наконец, кажется удивительным и маловероятным, чтобы случайная неоптимизированная последовательность вдруг стала кодировать нетоксичный белок, который быстро приобрёл бы функциональность и избежал бы давления отбора. Тем не менее, последние экспериментальные исследования показали, что это возможно.
Zhang et al. сравнивали 13 новых геномов, которые были недавно секвенированы и аннотированы из 11 близкородственных видов рисовых растений, включая наиболее распространенную пищевую культуру Oryza sativa (рис посевной, рис. 1).

Рис. 1. Рис посевной (Oryza sativa).
Чтобы не упустить из виду гомологичные некодирующие транскрипты предковых копий новых генов (ведь некодирующая ДНК быстро дивергирует до неузнаваемости), ученые исследовали эволюционно молодые виды Oryza, видообразование которых произошло в диапазоне от 0,4 до 14,3 миллиона лет назад. Исследуя эволюционную историю некодирующих транскриптов, они обнаружили не менее 175 генов, возникших de novo. Таким образом, авторы подтвердили возможность возникновения генов de novo и показали, что этот процесс характерен для всех эукариот (рис. 2), поскольку похожие данные были получены ранее на дрозофиле и млекопитающих.
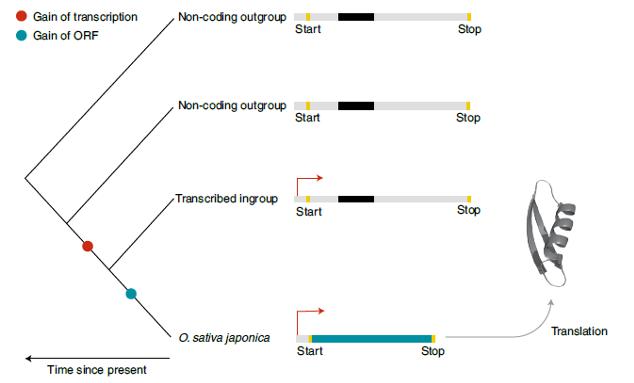
Рис. 2. Этапы возникновения гена de novo. Частота возникновения de novo генов риса по Zhang et al. составила 51,5 гена на миллион лет. Авторы реконструировали процесс возникновения этих генов. Они показали, что обычно появлению открытой рамки считывания (синий кружок на филогенетическом древе) предшествует усиление транскрипции (красный кружок на филогенетическом древе). На данном примере возникновению de novo специфичного для риса гена способствовала короткая делеция (выявлена при сравнении синтенных областей близкородственных видов), которая приблизила стартовый кодон и стоп-кодон к открытой рамке считывания, в результате чего у Oryza sativa (подвид japonica) сформировалась интактная ORF (синий цвет последовательности). Логично, что после транскрипции этой ORF её мРНК будет транслироваться в клетке, что приведет к появлению белковых продуктов, способных свободно приобретать новые функциональные возможности и стимулировать адаптационные процессы. Важно отметить, что Zhang et al. подтвердили трансляцию многих рисовых генов, возникших de novo, с помощью масс-спектроскопии. Они нашли доказательства того, что 57 процентов этих генов фактически транслируются в новые белки, формируя пул, состоящий из более 300 новых пептидов. Рисунок взят из статьи Bornberg-Bauer, Heames. Becoming a de novo gene // Nature Ecology & Evolution. 2019. V.3. Р. 524–525.
Используя данные по высокопроизводительному секвенированию РНК (high-depth RNA-seq) для изучения сравнительной экспрессии генов, Zhang et al. также смогли показать, что новые транскрипты и ORF возникают в геноме независимо (т.е. не являются следствием дупликации или рекомбинации), причем первые с гораздо большей скоростью (рис. 3а).
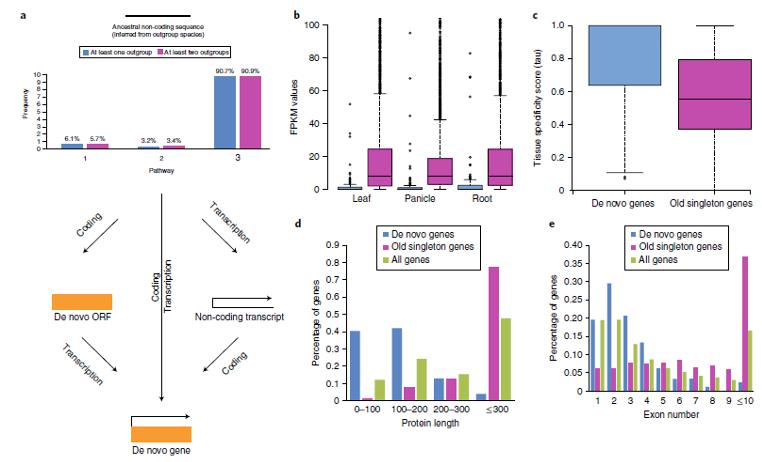
Рис. 3. Картины возникновения генов de novo, их экспрессии и структурных особенностей. а, Три пути возникновения гена de novo - анализ частоты возникновения из предполагаемых наследуемых некодирующих последовательностей 175 генов de novo: (1) модель ‘ORF-first’ (на первом этапе мутации превращали некодирующую последовательность в новую ORF, затем приобретённые регуляторные элементы (промоторы и энхансеры) активируют новую ORF в новый ген); (2) модель «транскрипция и ORF одновременно»; и (3) модель ‘transcription-first’. b, c, Паттерны экспрессии: новые гены (синие) имеют пониженную экспрессию (b) и более тканеспецифичны (c) по сравнению со старыми единичными генами (фиолетовые). Для определения значимости использовали критерий суммы рангов Уилкоксона с коррекцией на непрерывность (лист: P <0,0001; метелка: P <0,0001; корень: P <0,0001 на b; P <0,0001 на c). d, По сравнению со старыми единичными генами или всеми комбинациями генов, de novo гены короче. e, По сравнению со старыми единичными генами генам de novo свойственно иметь меньше экзонов, но наиболее распространённой их структурой является структура с двумя экзонами. Рисунок взят из статьи Zhang et al. Rapid evolution of protein diversity by de novo origination in Oryza // Nature Ecology & Evolution. 2019. V.3. P. 679–690.
Таким образом, авторы статьи получили доказательства в пользу модели, объясняющей появления генов de novo из некодирующей РНК (модель ‘transcription-first’). Большинство новых последовательностей дивергируют и теряются, однако, некоторые (10%) начинают стабильно транслироваться и сохраняются без существенного изменения свойств их белковых продуктов, т.е. поддерживаются естественным отбором. С другой стороны, пока неизвестно, что делают те белки, которые закодированы в этих рисовых генах. Авторы работы говорят, что собираются исследовать их в самое ближайшее время, но одновременно отмечают, что большинство генов de novo пока ещё не слишком почувствовали давление отбора. Те мутации, которые в них происходят, по тем или иным причинам не влияют (или не очень сильно влияют) на шансы растения выжить и оставить потомство; возможно, какой-то эволюционный вердикт им ещё предстоит в будущем.
Мы же отметим, что интересно также проследить, как в ходе эволюции меняется структура белков, кодируемых генами de novo. Это поможет понять, каким образом случайные изменения в их последовательности могут оптимизировать их функции. Таким образом, изучение генов, возникающих de novo, и их продуктов важно не только для понимания их функционального и биологического вклада в видообразование; они также несут информацию и расширяют наши представления об адаптивном направлении вектора эволюции генома.
Новость подготовила © Симонова О.Б.
18.11.2019