Комментарий к статье: Moroz LL, Nikitin MA, Poli ar PG, Kohn AB, Romanova DY. Evolution of glutamatergic signaling and synapses. Neuropharmacology. 2021 Jul 31:108740. doi: 10.1016/j.neuropharm.2021.108740. Epub ahead of print. PMID: 34343611
 © Дьяконова Варвара Евгеньевна, доктор биологических наук, профессор РАН. Главный научный сотрудник лаборатории нейробиологии развития.
© Дьяконова Варвара Евгеньевна, доктор биологических наук, профессор РАН. Главный научный сотрудник лаборатории нейробиологии развития.
Область научных интересов: нейроэтология, нейробиология, нейротрансмиттеры, поведение.
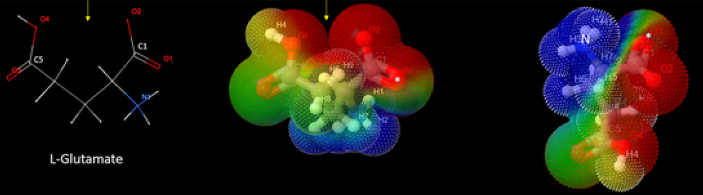
Идея о том, что глутамат мог быть первым трансмиттером, повышение концентрации которого в межклеточной среде сигнализировало о повреждении клетки, была озвучена Леонидом Морозом уже более 10 лет назад. Тогда она казалась интересной, но трудно доказуемой, интуитивной. Однако тем, кто прочитает монументальную статью 2021 года, посвященную эволюции глутаматергической сигнализации, эта идея уже не покажется оторванной от реальности. Годы интенсивной работы принесли замечательные результаты и далеко идущие выводы. Статьи такого масштаба (теоретического и экспериментального) - единичны.
Пару вводных слов о самом глутамате. Несколько причин ставят его в особенное положение по сравнению с другими сигнальными молекулами и, соответственно, служат источниками для интересных вопросов. Во-первых, большинство нейронов головного мозга млекопитающих использует глутамат, хотя существует более 20 других нейротрансмиттеров с низким молекулярным весом. Неудивительно, что около половины современных нейробиологов работают с различными аспектами глутаматергической передачи. Во-вторых, сравнительные исследования указывают на глутамат как на один из самых ранних трансмиттеров: глутаматные рецепторы выявлены группой Мороза и другими авторами практически у всех исследованных на сегодняшний день многоклеточных, включая растения. В-третьих, те же сравнительные исследования показывают, что глутамат, заняв лидирующее положение в мозге млекопитающих, по-видимому, утратил у них ряд функций и рецепторов, разнообразие которых у первичноротых выше. Например, у беспозвоночных глутамат часто действует и как тормозный нейротрансмиттер (Cleland, 1996; Kehoe et al., 2009; Kehoe, 2000) с использованием пентамерных рецепторных каналов с cys-петлями (Cymes, Grosman, 2021; Jaiteh et al., 2016; Kehoe et al., 2009; Lynagh et al., 2015). L-глутамат является нервно-мышечным нейротрансмиттером у большинства животных на планете, но не у млекопитающих (у которых эту функцию выполняет, как известно, ацетилхолин). Глутаматергические синапсы у моллюсков молекулярно более сложны, чем в мозге человека.
Наконец, в отличие от моноаминов и ацетилхолина, открытых в качестве нейротрансмиттеров существенно раньше, глутамат настолько распространенный клеточный метаболит, что исследователи долго ставили под сомнение саму возможность выполнения им трансмиттерной функции из-за его низкой специфичности и отсутствия специализированной системы синтеза, как у других трансмиттерных систем. Глутаматергический нейрон обладает эргичностью, то есть трансмиттерной специфичностью, в несколько ином смысле, чем, например, моноаминовые, петидергические или ацетилхолиновые нейроны. Если для последних характерна специализированная система ферментов синтеза, везикулярного захвата и выброса, специфичная только для этого типа клеток, то глутаматный нейрон становится таковым только за счет экспрессии гена глутаматного транспортера, закачивающего глутамат в везикулы – синаптические пузырьки. Стоп! Не здесь ли ниточка, потянув за которую, можно распутать большой клубок вопросов? Например, (i) почему и как получил глутамат универсальное применение во всех живых организмах? (ii) почему и как глутамат был задействован в качестве межклеточного сигнала? И (iii) почему и как глутамат стал доминирующим нейротрансмиттером в нашем мозге?
Авторы обсуждаемой статьи считают, что да. Что начинать разгадывать интересную задачку о происхождении трансмиттерной функции глутамата и ее дальнейшей эволюции нужно с рассмотрения положения этой аминокислоты во внутриклеточном метаболизме клетки.
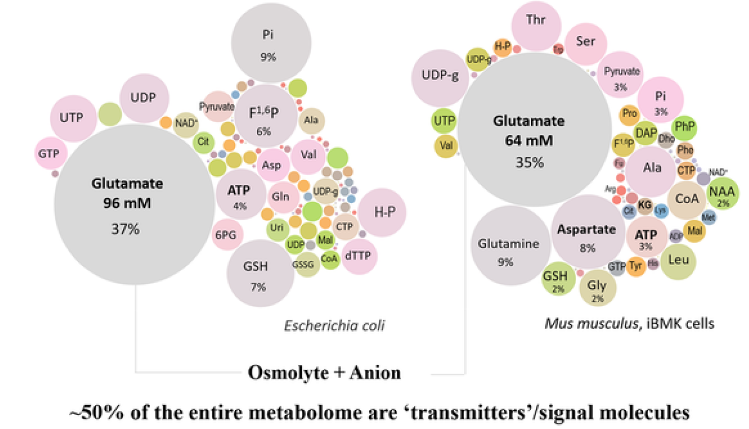
Простой рисунок (1) очень красноречив. Глутамат - самый распространенный клеточный метаболит. На рисунке показано соотношение разных внутриклеточных метаболитов у клетки бактерии и эпителиальной клетки мыши.
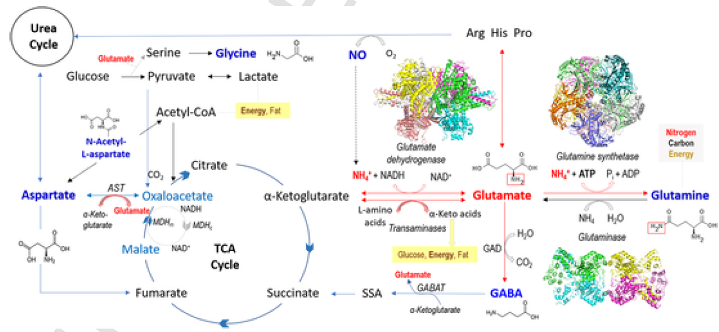
Следующая, более сложная схема (Рис. 2), хотя и не объясняет полностью, почему глутамата так много в клетках, хорошо иллюстрирует его участие сразу в нескольких базовых метаболических путях клетки. Как справедливо названа это часть статьи, глутамат - на перекрестке клеточного метаболизма.
Очевидно, что нарушение целостности любой клетки приведет к появлению в межклеточном пространстве самых разных метаболитов, но другим клеткам для выявления такого события проще настроиться на самый сильный и универсальный сигнал. Глутамат может смело претендовать на эту роль. Специфический химический сигнал о слабеющей или погибающей клетке может быть важным пищевым стимулом в одноклеточном мире. У многоклеточных повышение глутамата в межклеточнике становится уже сигналом, указывающим на неблагоприятные события в организме. Биологическая значимость этой информации очень высока, поэтому неудивительно, что уже на самых ранних этапах эволюции, по оценкам Л. Мороза с соавторами, 3.5 миллиарда лет назад, появляются системы распознавания экстраклеточного глутаматного сигнала, а именно глутаматные рецепторы. (Здесь трудно удержаться и не напомнить, что обе анцестральные функции глутамата, выявленные еще у донервных многоклеточных организмов Placozoa, несмотря на долгую полную событий эволюцию, прекрасно сохранились и у млекопитающих! Достаточно вспомнить про хорошо известный усилитель вкуса, глутамат натрия, взаимодействующий со специализированными вкусовыми рецепторами Умами, 旨味. )
Анализу эволюции генов глутаматных рецепторов посвящена значительная часть работы Л. Мороза с коллегами. Предполагается, что различные типы анцестральных глутаматных рецепторов у общего предка многоклеточных (Урметазоа, Urmetazoa) могли активироваться разными лигандами, включая глицин, глутамат и другие аминокислоты, как это показано, например, у современных растений и прокариот. Другими словами, эти рецепторы исходно могли быть универсальными аминокислотными сенсорами, и отчасти эта их функция сохранилась и у многоклеточных животных. Однако, относительно высокие концентрации высвобожденного внеклеточного глутамата (по сравнению с другими аминокислотами) могли сделать эти рецепторы функциональными рецепторами глутамата - аналогично iGluR, описанным у растений.
Расширенная филогения предполагает, что одним из старейших глутаматных рецепторов среди эукариот могут быть лямбда-рецепторы. Примечательно, что они были идентифицированы только у губок (Porifera). Пока неизвестно, оказались ли эти рецепторы инноваций губок либо потерялись со временем в других таксонах животных. Рецепторы NMDA-типа, хотя (к удивлению физиологов вышей нервной деятельности) выявлены у достаточно примитивных беспозвоночных, не были обнаружены вне многоклеточных животных. Среди метазоа, они также не обнаружены у гребневиков и губок. А всего у эукариот были идентифицированы двадцать две различные филетические линии глутаматных рецепторов (Рис. 3).
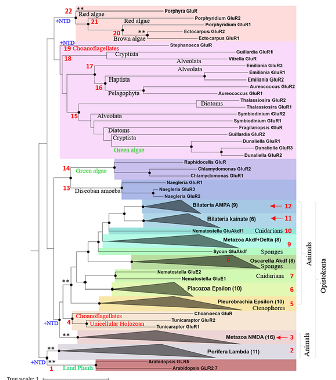
То, что на сегодняшний день известно о разнообразии глутаматергических систем многоклеточных, включая растения и водоросли, числе рецепторов и транспортеров глутамата, а также сигнальных функциях представлено на следующем рисунке (4).
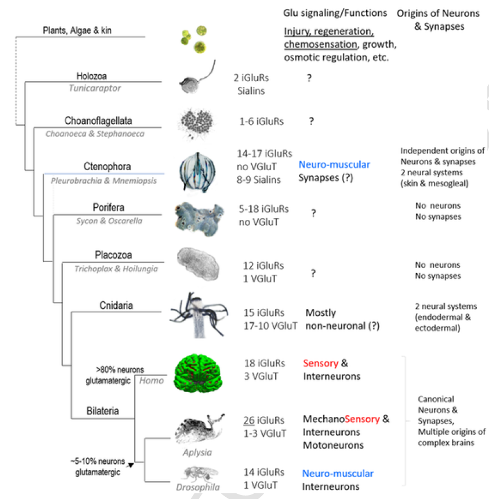
Существенные отличия видны даже в группе билатерий. У первичноротых только 5-10% нейронов использует глутамат в качестве нейротрансмиттера, в отличие от 50-80% нейронов в ЦНС млекопитающих. У последних глутамат уже не используется в качестве нейромышечного трансмиттера, а характерен для сенсорных систем и интернейронов. Почему мозг млекопитающих предпочел глутамат в качестве основного возбуждающего нейротрансмиттера? Авторы отвечают на этот вопрос так: очень низкая энергетическая стоимость производства (энергия, расходуемая на захват и синтез глутамата, у грызунов составляет всего 2–3% от общих затрат, связанных с передачей сигналов, 5% - у людей; кроме того, глутамат сам может быть источником энергии) и его простой запуск (все, что требуется для того, чтобы сделать глутаматергический нейрон – это активация экспрессии везикулярного глутаматного транспортера).
В дополнение к идее Л. Мороза можно предположить, что распространенность глутаматергических нейронов в мозге млекопитающих отражает «поспешность» его эволюции: объем наращивался быстро, простыми и максимально экономными способами. Если это действительно так, то кошмар химической школы Коштоянца-Сахарова (повышение химической однородности нейронов в мозге наиболее продвинутых в когнитивном отношении организмов противоречит основному постулату школы о принципиальной значимости для работы нервной системы мультитрансмиттерности и химического разнообразия), похоже, близок к своему разрешению. Эволюция мозга не уменьшала химическое разнообразие нейронов, она лишь была вынуждена на определенном этапе стремительно увеличить число нейронов, и сделала это самым простым, быстрым и дешевым способом.
В ответе Л. Мороза на вопрос о том, почему глутамат был выбран в качестве основного нейротрансмиттера млекопитающих, как мы видели, есть два объяснения: быстро и дешево. А вот от того, какое из них важнее, по-видимому, зависит дальнейшая эволюция нервной системы. Если нет серьезных энергетических ограничений, то быстрое увеличение числа нейронов, выполненное наиболее простым способом, таит в себе огромный потенциал для будущей эволюции, но уже не за счет числа, а за счет увеличения химической разнородности нейронов.
В заключение, настойчиво рекомендую прочитать исходную статью целиком, поскольку я пересказала лишь небольшую её часть.
Новость подготовила © Дьяконова В.Е.
19.08.2021