 © Симонова Ольга Борисовна, руководитель лаборатории МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИХ ПРОЦЕССОВ РАЗВИТИЯ, доктор биологических наук.
© Симонова Ольга Борисовна, руководитель лаборатории МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИХ ПРОЦЕССОВ РАЗВИТИЯ, доктор биологических наук.
Яйцекладущие – самый древний и примитивный отряд млекопитающих. Они, как и остальные млекопитающие, ведут происхождение от рептилий, однако отделились от них достаточно рано, образовав собственную ветвь развития. Яйцекладущие сочетают в себе черты древних рептилий, птиц и звероящеров. В частности, представители отряда откладывают яйца, у них есть клюв, но при этом выкармливают потомство молоком. Этот подкласс представлен единственным отрядом под названием однопроходные, который в свою очередь объединяет два семейства: ехидновые и утконосовые. Эти уникальные животные обладают рядом особенностей: у них нет сосков - детеныши слизывают молоко, которое выделяется из трубчатых желез, прямо с шерсти матери. Они довольно ядовиты. Имеют специальную клоаку, в которую открываются и мочевой пузырь, и кишечник. Помимо этого, утконосы и ехидны являются безголосыми и беззубыми. Учёные под руководством профессора Аделаидского университета (Австралия) Франка Грутцнера проанализировали геном утконоса и ехидны и выяснили, как они стали одними из самых странных животных на Земле.
В общей сложности генетики выделили более 20 и 22 тысяч генов в ДНК утконосов и ехидн соответственно. Затем их распределили по хромосомам. Сопоставив наборы мелких мутаций и различия в устройстве генов у обоих животных, биологи пришли к выводу, что их предки разделились уже после вымирания динозавров, в начале эоцена, примерно 55 млн лет назад. Их общий предок отделился от предков плацентарных и сумчатых 187 млн лет назад, в первой половине юрского периода. Эти оценки, как отмечают ученые, хорошо согласуются с данными палеонтологов (Fig. 1).
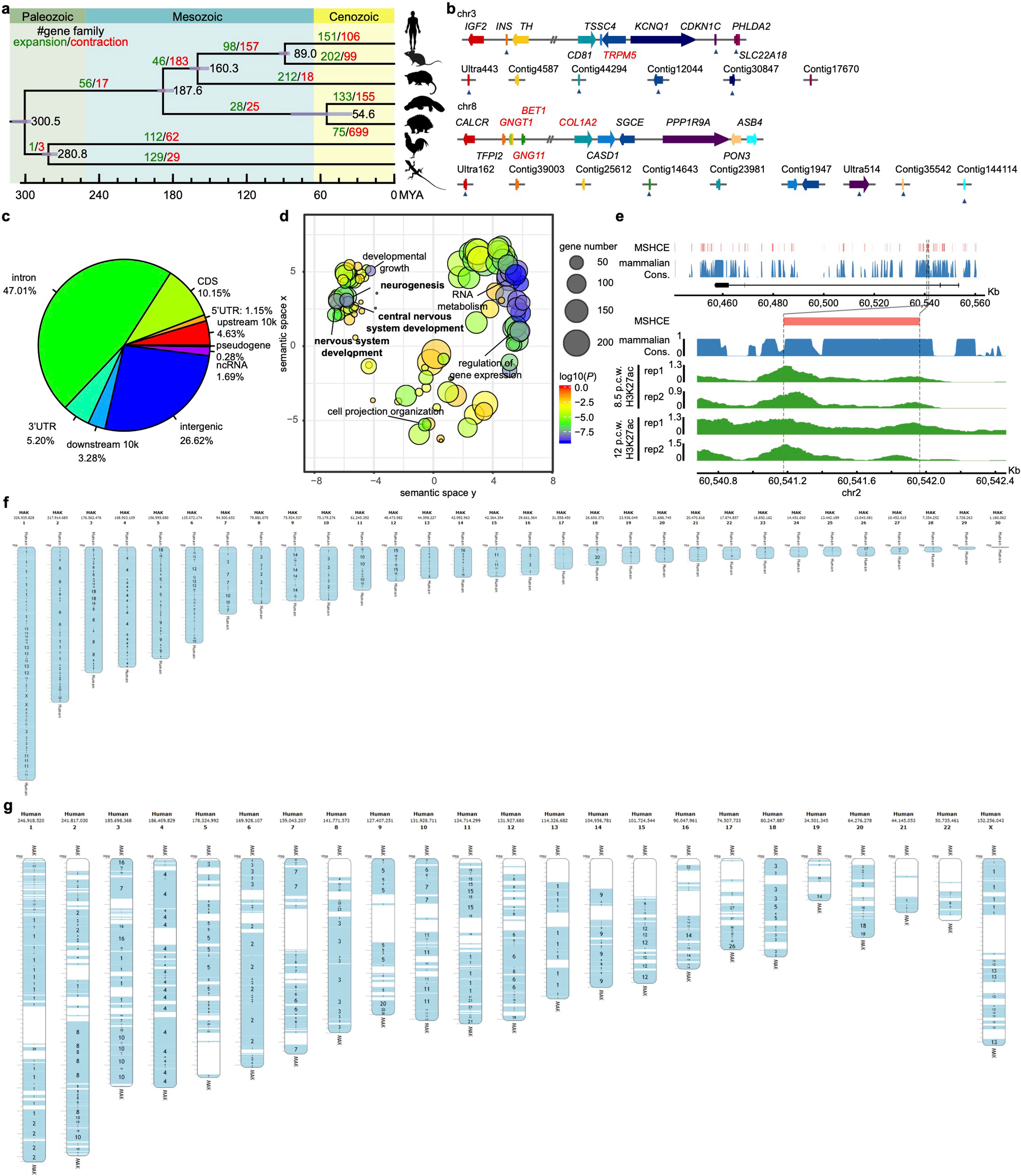
Fig. 1. Филогенетическое древо построено с использованием четырехкратно вырожденных сайтов из 7 946 ортологов один к одному среди семи репрезентативных видов (человек, мышь, опоссум, утконос, ехидна, курица и зеленая ящерица). Количество семейств генов, которые претерпели значительные распространения (зеленый) и сокращения (красный), отмечены на каждой ветви. Коррекция множественного тестирования не применялась.
Геном утконоса уже был частично расшифрован ещё в 2008 году (сборка генома OANA5). Благодаря его анализу ученые узнали, что у этих австралийских экзотов обилие половых хромосом: у утконоса имеется 10 половых хромосом, из которых 5 являются Х-хромосомами и 5 - Y-хромосомами, которые соединены друг с другом особым образом во время мейоза. Структура одной из "женских" хромосом оказалась похожа на Z-хромосому рептилий и птиц.
В ходе нового исследования генетики не просто полностью расшифровали геномы утконосов и ехидн, но и выяснили, каким образом он разбит на хромосомы и как они распределены внутри ядра их клеток (Fig. 2). Подобные сведения, по словам Грутцнера и его коллег, критически важны для изучения истории эволюции генома, в том числе и того, как возникли уникальные половые хромосомы утоконосов и ехидн.
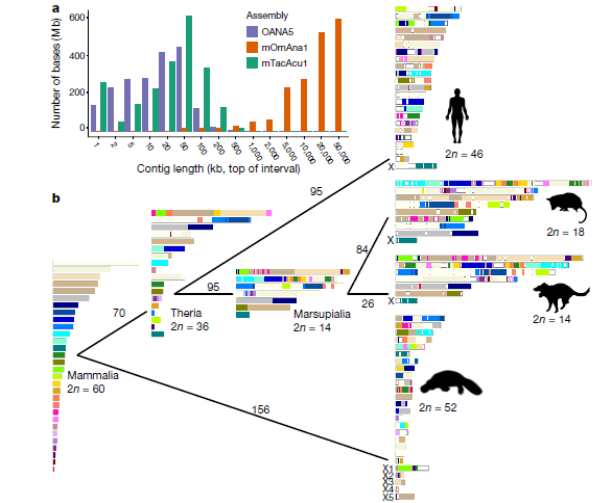
Fig. 2. Хромосомный набор в эволюции геномов яйцекладущих (monotreme) и остальных млекопитающих.
а - Распределение длин набора контигов у трех яйцекладущих показывает значительное улучшение качества сборки последовательностей с точки зрения её непрерывности у утконоса (зелёный) и, по крайней мере, адекватное качество у ехидны (оранжевый). Предыдущая сборка генома самки утконоса (OANA5 - фиолетовая) предоставила многие важные открытия в монотремной биологии и млекопитающих эволюция. Однако только около 25% его последовательности было отнесено к хромосоме 2. Неполная сборка генома утконоса без последовательностей Y-хромосом и отсутствие генома ехидны ограничили интерпретацию эволюции млекопитающих и монотрем.
b - Траектория эволюции кариотипа млекопитающих. 2n = 60 – предполагаемый кариотип общего предка млекопитающих. Консервативные блоки обозначены цветом в соответствии с их хромосомным положением в предке млекопитающего. Для каждой ветви показано количество предполагаемых перегруппировок.
Особый интерес представляют половые хромосомы яйцекладущих млекопитающих, которые возникли независимо от таковых у высших млекопитающих, за счет добавления аутосомы на предковую пару XY, в результате чего образуется система множественных половых хромосом, которая собирается в цепочку во время мейоза. Похожесть структуры и формы половых хромосом утконосов и ехидн показывает, что эта уникальная система появилась еще у их общего предка. Ученые предполагают, что изначально эти хромосомы были объединены в необычную кольцеобразную структуру, которая впоследствии распалась на связанные друг с другом части. В пользу этого, в частности, говорит то, что копии генов из последней "мужской" хромосомы Y5 есть и в первой "женской" хромосоме X1 (Fig. 3).
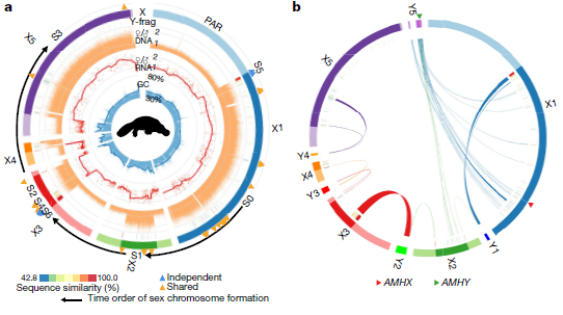
Fig. 3. Происхождение и эволюция половых хромосом утконоса.
а - Геномный состав половых хромосом утконоса. От внешнего кольца к внутреннему:
- Х-хромосомы с маркированными PAR - области, которые имеют общие последовательности между половыми хромосомами (светлые цвета), и участки SDR, которые дифференцировались по половому признаку (темные цвета);
- фрагменты, собранные в Y-хромосомы, в основном соответствуют участку SDR, величина расхождения с гомологичными последовательностями X-хромосом отмечена уровнем в цветовой шкале;
- соотношение самок и самцов (F/M) коэффициенты покрытия ридов короткого секвенирования в неперекрывающихся окнах размером 5 кб;
- соотношение экспрессии генов F/M (каждая красная точка - один ген) в ткани взрослой почки и тенденция к сглаживанию экспрессии; GC-контент в неперекрывающихся окнах размером 2 кб. Кроме того, на кольце Х-хромосомы авторы пометили позиции гаметологичных участков, которые подавляют рекомбинацию до расхождения яйцекладущих («общие», оранжевые треугольники) и после расхождения («независимые», синие треугольники).
b - Гомология между X и Y хромосомами утконоса. В частности, большая часть Y5 обнаруживает гомологию с X1 и X2, что указывает на конформацию предкового кольца половых хромосом утконоса. Обозначили положение предполагаемого гена, определяющего пол, AMH.
Напомним, что в мейозе гомологичные хромосомы объединяются и формируют биваленты. У человека Y-хромосома потеряла так много генов, что участков гомологии с X-хромосомой почти не осталось, поэтому рекомбинации между этими хромосомами практически не происходит. Но у многих видов различия между половыми хромосомами не столь значительны, и в мейозе они, как и аутосомы, соединяются с образованием бивалента. Однако, как ведут себя при мейозе несколько половых хромосом? У самцов, имеющих несколько Х-хромосом и несколько Y-хромосом, образуются не биваленты, а особые кольцевые структуры, в которых каждая хромосома взаимодействует с гомологичными участками двух своих соседей. Это возможно благодаря тому, что на всех X- и Y-хромосомах имеются гомологичные псевдоаутосомные районы. На финальных этапах мейоза кольцевая структура упорядоченно разделяется, и сперматозоиды получают либо n Х-хромосом, либо n Y-хромосом, где n — число хромосом в гаплоидном хромосомном наборе данного вида. Следовательно, наличие характерной кольцевой структуры в мейозе клеток-предшественников сперматозоидов является верным признаком наличия у вида нескольких половых хромосом.
Грутцнер и его коллеги объясняют, что раньше кольцеобразные объединения хромосом никогда не находили среди животных – их встречали только у растений. Однако справедливости ради отметим, что именно такую структуру удалось обнаружить в сперматоцитах лягушки - пятипалого свистуна - с помощью флуоресцентной микроскопии в 2018 г. (Fig. 4).
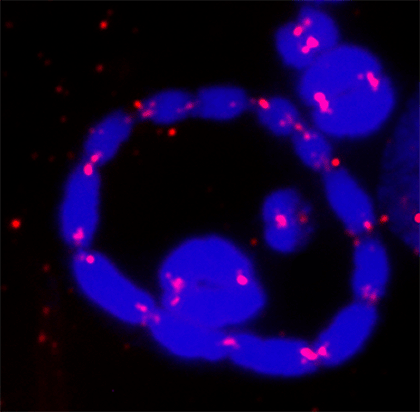
Fig. 4. Кольцевая структура, формирующаяся при мейозе у самцов пятипалого свистуна. Отчетливо видны 12 хромосом, образующих кольцо. ДНК окрашена синим, красным выделены теломеры. Картинка взята из T. Gazoni et al., (2018). More sex chromosomes than autosomes in the Amazonian frog Leptodactylus pentadactylus. Chromosoma. 127, 269-278.
В рассматриваемой работе ученые провели детальный анализ контактов половых хромосом, используя технологию Hi-C (High conformation Capture –"конформации хромосом высокого порядка"), которая позволяет определять все хромосомные контакты в ядре клетки) (Fig. 5). Они надеются, что дальнейшее изучение геномов утконосов и ехидн поможет им понять, как возникла эта конфигурация хромосом.
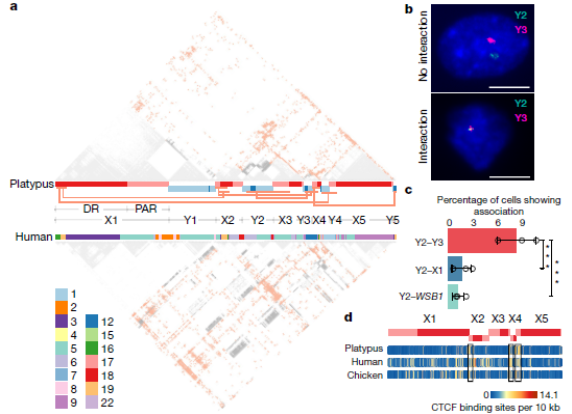
Fig. 5. Взаимодействие половых хромосом утконоса.
а - Межхромосомные взаимодействия между половыми хромосомами утконоса, обнаруженные технологией Hi-C (High conformation Capture –"конформации хромосом высокого порядка" позволяет определять все хромосомные контакты в ядре клетки) в ткани печени утконоса (вверху) и человека (внизу). Полосы между панелями Hi-C показывают половые хромосомы утконоса и их ортологи в геноме человека. Серый, внутрихромосомные взаимодействия; красный, межхромосомные взаимодействия. Красные линии связывают области со значительно высокими межхромосомными взаимодействиями. Межхромосомные взаимодействия, по-видимому, сохраняются у млекопитающих, на что указывают гомологичные хромосомные фрагменты половых хромосом человека и утконоса и их паттерны контактов на Hi-C.
b - FISH с зондами BAC для обнаружения половых хромосом Y2, Y3 или X1 и аутосомной хромосомы 17 (WSB1) в интерфазных фибробластах утконоса. Примеры показывают отсутствие взаимодействия между хромосомами Y2 и Y3 (вверху, n = 593, 3 независимых эксперимента) и их взаимодействие (внизу, n = 56, 3 независимых эксперимента). Масштабные полоски - 10 мкм.
c - значительно более высокая частота взаимодействия между Y2 и Y3, чем между Y2 и X1, а также между Y2 и WSB1 (хромосома 17).
d - Плотность предполагаемых сайтов связывания с белком CTCF (белок-инсулятор, контролирует границы активного хроматина), показывающая его обогащение среди гомологичных областей в геномах утконоса, человека и курицы.
Утконосы и ехидны радикально различаются в питании (плотоядные по сравнению с насекомоядными), сенсорной нейрофизиологии (ориентированные на электрорецепцию по сравнению с ориентированными на обоняние), а также в специфических внутривидовых отношениях и защитных адаптациях. Благодаря своим отчетливым экологическим, анатомическим и физиологическим особенностям, монотремы представляют собой млекопитающих, хорошо подходящих для изучения эволюции экологической адаптации. Например, у млекопитающих имеется более 25 копий генов рецепторов горького вкуса (гены TAS2R), тогда как это семейство генов значительно меньше у монотремов, только 7 у утконоса, у ехидны это число сокращается до трех (Fig. 6а). Подобное сокращение наблюдается у панголинов с похожей диетой (насекомоядные), говорящее о конвергентной эволюции. Исследователи надеются, что дальнейшее изучение геномов утконосов и ехидн поможет им понять, как яйцекладущие "потеряли" гены, которые отвечают за распознавание горького вкуса.
Как и у утконоса, «клюв» ехидны хорошо иннервирован. В его коже расположены как механорецепторы, так и особые клетки-электрорецепторы; с их помощью ехидна улавливает слабые колебания электрического поля, возникающие при движении мелких животных. Ни у одного млекопитающего, помимо ехидн и утконоса, подобного органа электролокации не обнаружено. Носовая полость утконоса закрывается во время ныряния, и размер основной обонятельной луковицы утконоса намного меньше, чем у ехидны. В соответствии с этим количество обонятельных рецепторов у утконоса (299) намного меньше, чем у ехидны (693) (гены OR - рис. 6a). Различия в размере обонятельной луковицы и обонятельных рецепторах OR у ехидны могут способствовать поиску по запаху подземной добычи, тогда как утконос полагается на электрорецепцию для обнаружения добычи в воде. Однако размеры добавочной обонятельной луковицы у утконоса больше, чем у ехидны. В дополнительную обонятельную луковицу идут проекции нейронов из вомероназального органа (периферический отдел дополнительной обонятельной системы некоторых позвоночных животных, который играет важную роль в формировании полового поведения), поэтому у утконоса наблюдается заметное увеличение количества вомероназальных рецепторов типа 1 (262) по сравнению с ехидной (28) (гены V1R - рис. 6a). «Вомероназальные рецепторы играют важную роль в ухаживании, заботе о потомстве, индукции лактации и выделении молока у однопроходных. Таким образом, диверсификация систем обонятельной луковицы и дополнительных обонятельных луковиц у яйцекладущих представляет собой интересный пример эко-эволюционного компромисса» - отмечают исследователи. Сильное развитие V1R связана с размером вомероназального органа и ночной активностью. Это также согласуется с тем фактом, что утконос закрывает глаза при нырянии и, следовательно, полностью полагается на другие чувства под водой и в норе.
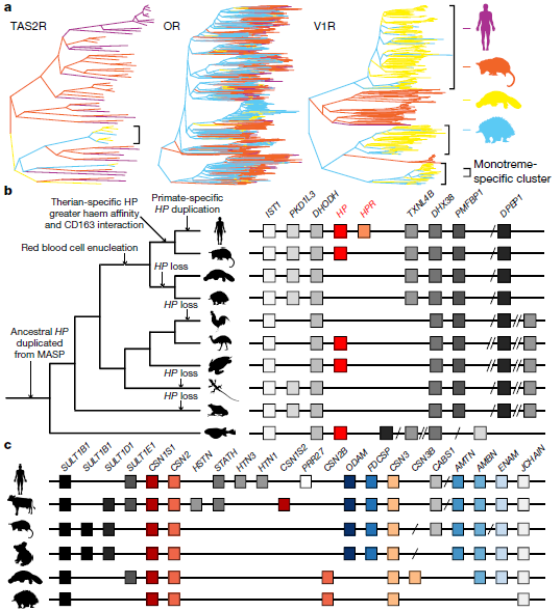
Fig. 6. Геномные особенности, связанные с биологическими характеристиками яйцекладущих.
а - Различия в количестве генов рецепторов горького вкуса TAS2R, и обоняния OR и V1R между утконосом и ехидной.
b - Филогения и синтения гена гаптоглобина HP. Регионы не масштабированы.
c - Сохранение синтении области, окружающей гены казеина (гены CSN - красные) и гены, определяющие зубы (ODAM, FDCSP, AMTN, AMBN и ENAM - голубые).
Деградация гемоглобина у монотрем
Полуводный образ жизни утконоса поддерживается особенно высоким уровнем гемоглобина и большим количеством мелких красных кровяных телец. Система детоксикации гемоглобин-гем у млекопитающих обеспечивает эффективный клиренс для минимизации окислительного повреждения. В эту систему входит гаптоглобин - шаперон гемоглобина, и гемопексин с микроглобулином альфа-1, с которыми связывается свободный гем. Интересно, что и гемопексин, и гены микроглобулина альфа-1 обнаружены в монотремных геномах, тогда как ген гаптоглобина отсутствует (Fig. 6b). Это говорит о том, что у монотремов эволюционировала другая система деградации гемоглобина. Некоторые позвоночные, не являющиеся млекопитающими, также потеряли ген гаптоглобина, включая курицу (Fig. 6b), но у них есть альтернативный секретируемый шаперон PIT54, связывающийся с гемоглобином.
Переход от яйцекладки к живорождению
Прежде всего, утконосы и ехидны – единственные современные млекопитающие, откладывающие яйца (поэтому их выделяют в подкласс яйцекладущих). И теперь понятно, как им это удаётся. У этих двух животных всё ещё работают гены, отвечающие за формирование вителлогенина – предшественника яичного желтка (VTG). Правда, не в полном составе. У общего предка млекопитающих и птиц таких генов было три. Птицы и рептилии сохранили все три функциональные копии, у монотремов авторы обнаружили только одну его функциональную копию (VTG2) (два других было потеряно примерно 130 миллионов лет назад). Человек и почти все остальные млекопитающие полностью утратили эти гены. Вместо выключившихся генов, отвечающих за вителлогенин, все млекопитающие приобрели гены синтеза казеина – основного белка молока. То же самое произошло с утконосом и ехидной, хотя у них были заменены не все "вителлогениновые" гены.
Подобно сумчатым, у яйцекладущих млекопитающих длительный период лактации, и состав молока меняется по мере развития, в соответствии с потребностями потомства. SPINT3, основной специфический для молока белок, который присутствует в ранней лактации млекопитающих и играет роль в иммунной защите потомства сумчатых животных. У однопроходных он отсутствует. Анализ синтении подтвердил, что эта область является консервативной у утконоса, но содержит две копии генов, кодирующих новые белки с доменом Kunitz. Kunitz–белки это быстро эволюционировавшее семейство, и один из его новых членов у монотремов имеет иммунопротекторную функцию, аналогичную SPINT3. В геноме монотрем, как и в геномах остальных млекопитающих есть множество генов, кодирующих белки молока. У большинства млекопитающих три казеиновых гена, которые кодируют секретируемые молочные белки (Fig. 6c). У монотрем есть дополнительные гены казеинов с неизвестными функциями: копия CSN2 (CSN2B) и копия CSN3 (CSN3B). Все казеины принадлежат семейству секреторных кальций-связывающих фосфопротеинов (SCPP) и, предполагается, что гены казеина произошли от гена ODAM, связанного с развитием зубов, через его предшественников FDCSP и SCPPPQ. Предок монотрем (яйцекладущих) имел зубы, но у обоих современных монотрем нет зубов. Ученые в этом исследовании показали, что из восьми генов, участвующих в развитии зубов, четыре гена были потеряны (Fig. 6с), что согласуется с данными по другим беззубым или лишенным эмали плацентарными животными. Ехидны (но не утконосы) в дальнейшем потеряли два гена эмали: ODAM, и FDCSP. Анализ синтении показал, что дополнительные гены казеина у монотерм (CSN2B и CSN3B) обнаружены в той же области хромосомы млекопитающих, что и ODAM и FDCSP, т.е. внутри локуса, кодирующего казеины (Fig. 6c), что является дополнительным доказательством того, что казеины произошли от одонтогенных генов, контролирующих развитие зубов.
В итоге авторы исследования заключили, что полные и точные аннотации геномов имеют решающее значение для эволюционного и функционального анализа. По-прежнему существует проблема их высокоточной сборки на уровне хромосом, особенно для дифференцированных половых хромосом. Генетики получили высококачественный геном утконоса, используя комбинацию технологии одномолекулярного секвенирования и нескольких методов физического картирования, чтобы как можно точнее собрать прочтённые последовательности на хромосомах. Авторы уделили особое внимание анализу древней сенсорной системы и специфике её изменений, деградации гемоглобина и репродуктивной функции, которые представляют самые интересные биологические особенности утконоса и ехидны. Новые геномы обоих видов позволят глубже понять приобретенные свойства плацентарных и сумчатых млекопитающих, а также биологию и эволюцию необычных млекопитающих, откладывающих яйца.
Новость подготовилa © Симонова О.Б.
19.01.2021