
Научные интересы: морфогенезы губок, цитоскелет, актомиозиновая сократимость, клеточная подвижность.
Форма клетки, её подвижность, безошибочное прохождение через чекпойнты клеточного цикла и способность отвечать на сигналы внешней среды зависят от систем цитоскелета и их регуляторов. Микротрубочки (МТ) – вторые по представленности (после актиновых филаментов) и крайне консервативные у эукариот элементы цитоскелета (Ludueńa et al., 1977). МТ, помимо их роли в формировании веретена деления, центриолей, аксонем ресничек и жгутиков, обеспечивают позиционирование органелл в клетке, транспорт везикул, клеточную подвижность и дифференцировку. В основе всех этих функций лежит «динамическая нестабильность» МТ: регуляция их длины за счёт быстрой смены процессов полимеризации и деполимеризации (Mitchison & Kirschner, 1984). Динамику МТ регулируют множество групп белков. Кроме того, в последние годы интерес исследователей привлёк «тубулиновый код» (Verhey & Gaertig, 2007). По аналогии с «гистоновым кодом», «тубулиновый код» — это пост-трансляционные модификации (ПТМ) тубулина, которые локально изменяют динамику МТ, обеспечивая выполнение специфических функций. ПТМ тубулина были найдены как у позвоночных, так и у беспозвоночных животных. Например, уровень ацетилирования α-тубулина определяет правильность формирования механосенсорных нейронов у Caenorhabditis elegans (Shida et al., 2010).
Возникает вопрос: изменялось ли строение тубулинового цитоскелета и механизмы его регуляции в ходе эволюции Metazoa? Ответить на него мог бы сравнительный анализ сетей МТ в клетках животных из филогенетических удалённых групп. Однако проблема состоит в том, что о системе МТ в клетках древнейших животных, губок (тип Porifera) имеются только фрагментарные данные. Известно, что MT у губок участвуют в позиционировании и транспорте органелл и везикул, а также реорганизуются во время миграции клеток (Kirfel & Stockem, 1997; Wachtmann & Stockem, 1992). При этом данных об архитектуре системы МТ, механизмах её регуляции и роли в морфогенетических процессах практически нет.
В статье, опубликованной в журнале Cell and Tissue Research, коллектив авторов не только описал архитектуру сетей микротрубочек в клетках известковой губки Leucosolenia corallorrhiza в норме и при регенерации, но и продемонстрировал распределение в клетках "тубулинового кода" (тирозинирования, детирозинирования и ацетилирования тубулина). Эти результаты, полученные на представителе базальной ветви древа Metazoa, позволяют ввести исследования тубулинового цитоскелета и регуляции его работы в эволюционный контекст.
В качестве модели использовались интактные особи Leucosolenia, а также кольцевидные фрагменты тела губки. На торцевых поверхностях получившихся фрагментов формируется регенеративная мембрана (РМ), образованная клетками, закрывающими раневую поверхность (Рис. 1). На этой модели очень удобно отслеживать поведение клеток при регенерации, а также исследовать их цитоскелет методами иммуноцитохимии и конфокальной микроскопии. РМ, как и интактная стенка тела, состоит из трёх клеточных слоев: наружного (экзопинакодемы), внутреннего (хоанодермы) и лежащего между ними мезохила – системы из межклеточного матрикса, минеральных скелетных элементов (спикул) и одиночных свободно мигрирующих клеток (Lavrov et al., 2018).
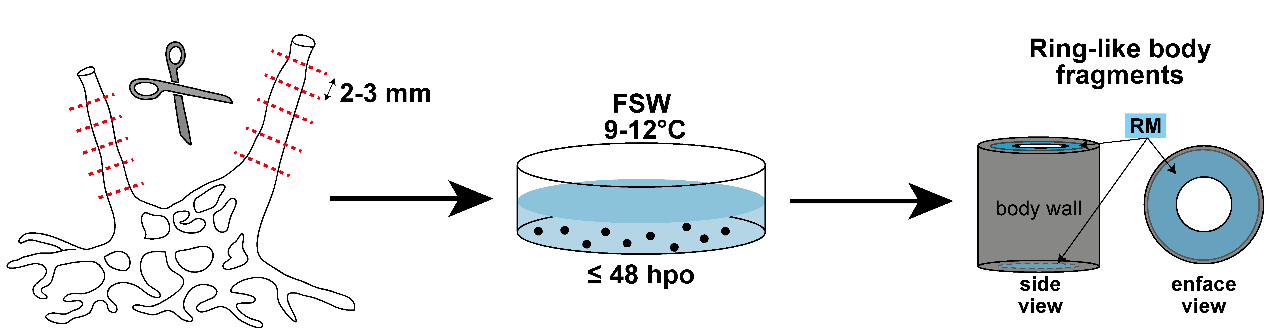
Рисунок 1. Схема эксперимента, иллюстрирующая модель получения кольцевидных фрагментов тела. FSW – фильтрованная морская вода; hpo – часы после операции; RM – регенеративная мембрана.
Оказалось, что сети микротрубочек хорошо выявляются у губки во всех типах клеток. В экзопинакоцитах (покровных клетках) интактной губки обнаружилась сеть МТ, имеющая один ЦОМТ вблизи ядра и состоящая из искривлённых пучков МТ, направленных от этого ЦОМТа в цитоплазматическую пластинку, где они расходятся радиально (Рис. 2, верхняя панель). Интересно также, что клеточные выросты экзопинакоцитов имеют тубулиновый скелет (в дополнение к актиновому). Авторы предполагают, что такие выросты, соединяясь друг с другом, позволяют клеткам обмениваться информацией, что особенно важно для губок в условиях отсутствия у них нейрогуморальной регуляции (Рис. 2, нижняя панель).
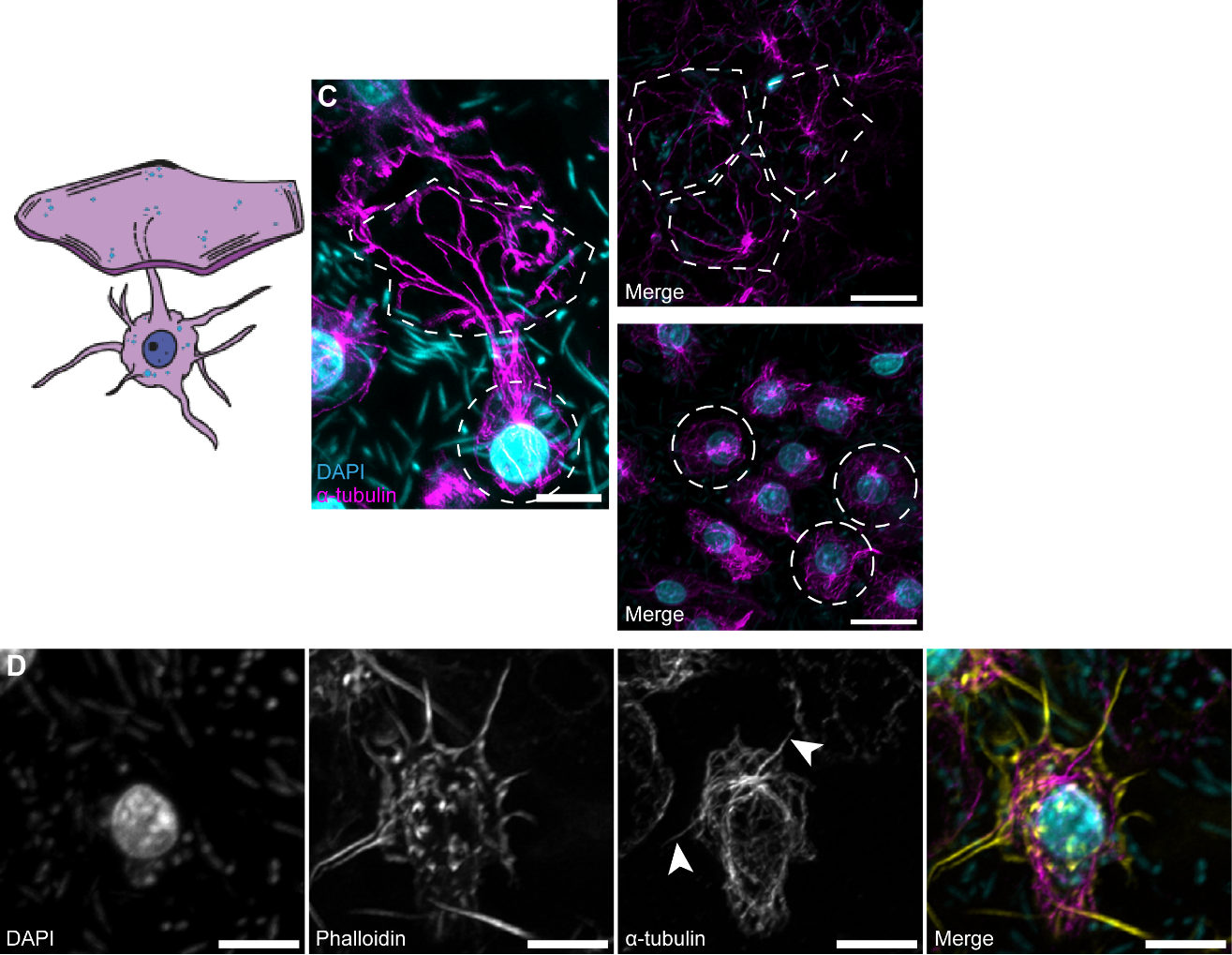
Рисунок 2. Системы цитоскелета экзопинакоцита интактной губки. Верхняя панель: система МТ экзопинакоцита, схема клетки: полигональная цитоплазматическая пластинка и непосредственно тело клетки с выростами. Нижняя панель: выросты экзопинакоцита с актиновым и тубулиновым кором (белые наконечники стрелок). КЛСМ, максимальные проекции интенсивности.
В экзопинакоцитах "тубулиновый код" представлен ацетилированным тубулином в веретенах деления, а также детирозинированным тубулином, ассоциированным с центриолями или перицентриолярным материалом. Оказалось, что такое распределение тирозинированного тубулина характерно и для других типов клеток Leucosolenia, причем как в интактных клетках, так и при регенерации.
В экзопинакоцитах, образующих РМ (т.е. в эксперименте по регенерации), архитектура сети МТ не меняется. Однако у клеток лидирующего края РМ обнаружена интересная особенность: часть пучков МТ лежит параллельно краю РМ. По мнению авторов, это может быть связано с сильным механическим напряжением, направленным по касательной к краю раны. Такое распределение механических сил скорее всего обусловлено сокращением актомиозинового тяжа, опоясывающего рану (Skorentseva et al., 2023).
В статье впервые подробно охарактеризована архитектура сети МТ в хоаноцитах – уникальных клетках губок, обладающих воротничком и жгутиком и составляющих основу водоносной системы. Во-первых, в хоаноцитах была обнаружена сеть МТ, считающаяся типичной для призматических эпителиев (напомним, что эпителии губок нельзя отнести к «классическим»). Для такой сети МТ характерна ярко выраженная полярность архитектуры: пучки МТ идут из апикальной части клетки в базальную, без выраженного единого ЦОМТа (Рис. 3). Хотелось бы отметить две интересных особенности организации этой сети у Leucosolenia.
(1) Под апикальной поверхностью клетки обнаружена структура, которую авторы статьи назвали «колесом со спицами» (spoke wheel). В плоскости апикальной поверхности клетки пучки микротрубочек расходятся радиально из центра, а затем изгибаются в направлении базального конца клетки, и идут параллельно друг другу вдоль апикобазальной оси клетки (Рис. 3). В области базальной поверхности клетки сеть микротрубочек плотная, состоит из коротких искривлённых пучков, без единого ЦОМТа и сложно разрешимая оптически.
(2) Другая интересная особенность связана с "тубулиновым кодом". Ацетилированные МТ, вероятно входившие в состав веретена деления, составляют основу цитоскелета недавно прошедших митоз дочерних хоаноцитов (Рис. 4), а не только жгутиков в этих клетках. Детирозинированных МТ в хоаноцитах обнаружено не было.
По мнению авторов, поляризованная сеть МТ в хоаноцитах губок обеспечивает транспорт вакуолей с частицами пищи, которые хоаноциты поглощают и передают другим клеткам.
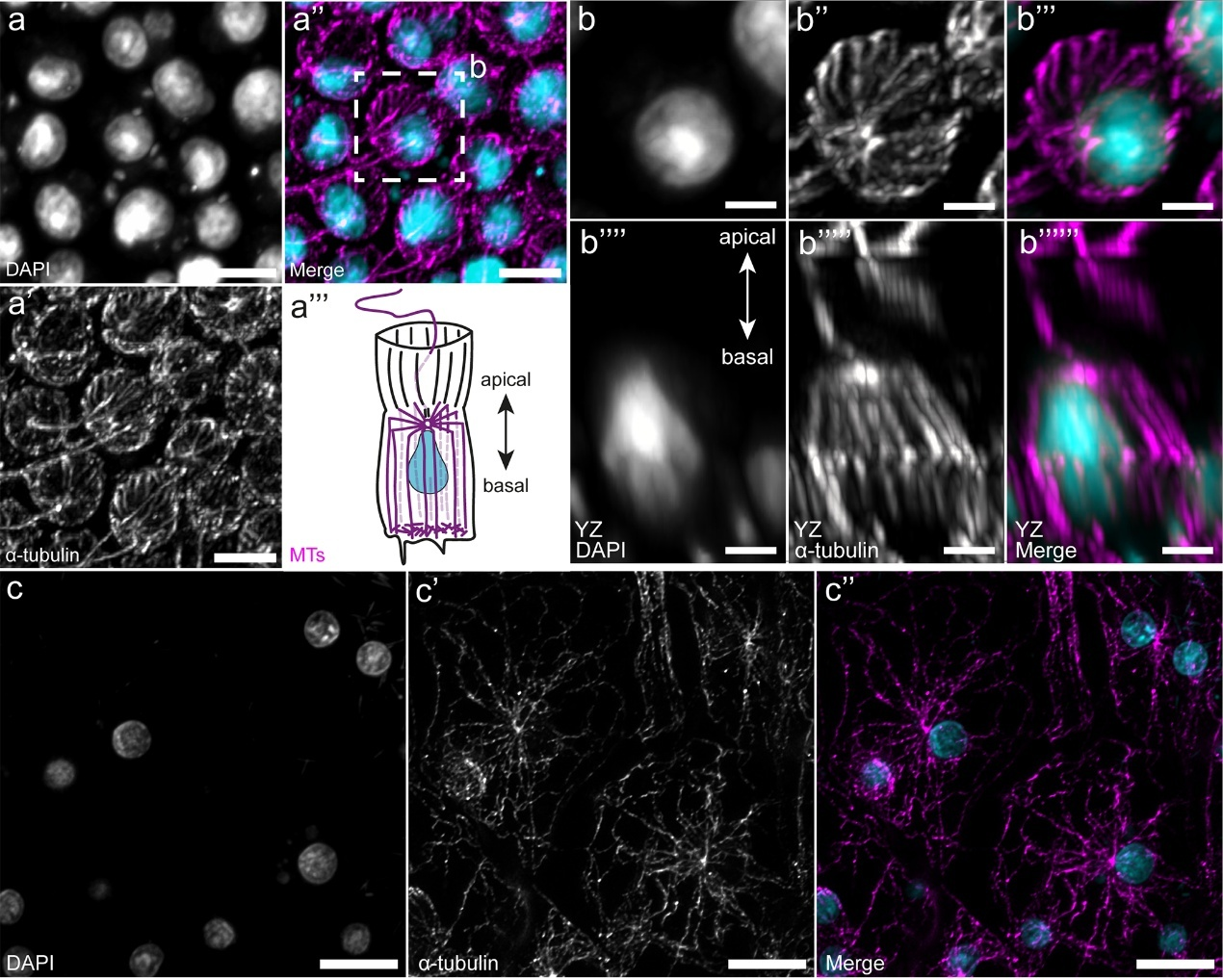
Рисунок 3. Микротрубочки в хоаноцитах интактной губки. (a - a´´) «Колесо со спицами» (“spoke wheel”) в плоскости апикальной поверхности хоаноцита, (а''') - схема хоаноцита. (b) Увеличена область, обведенная рамкой на (a´´), (b - b´´´´´´) - разные ортогональные проекции. КЛСМ, максимальные проекции интенсивности. Масштабный отрезок: (a) 5 мкм, (b) 2 мкм.
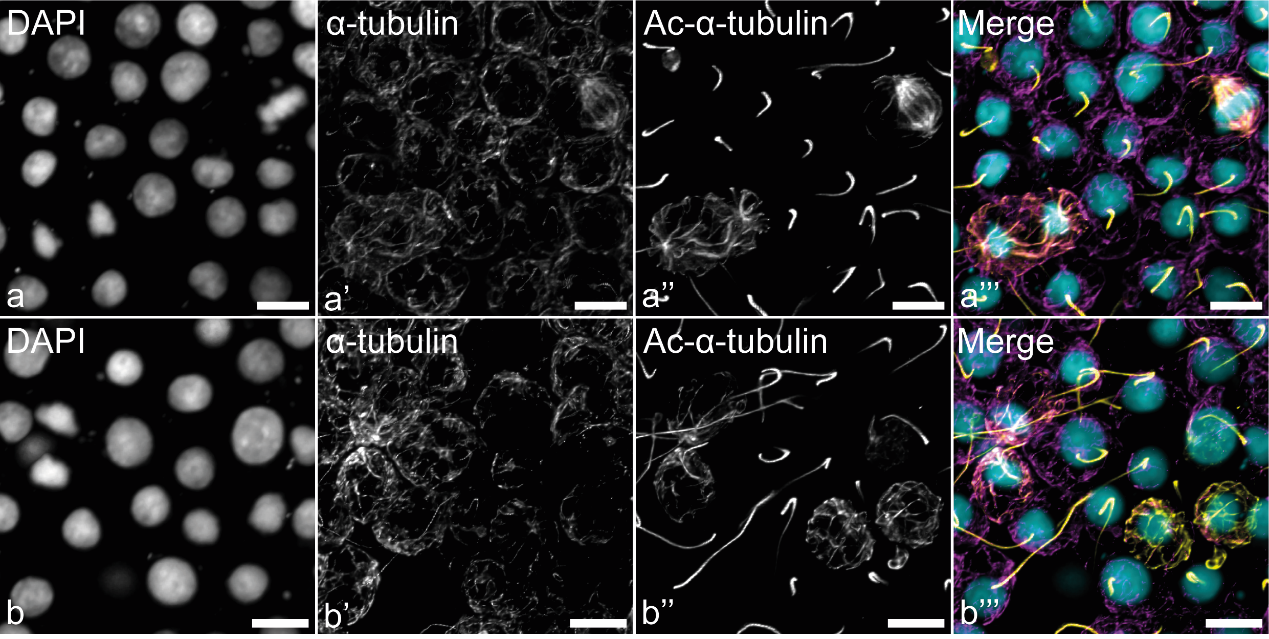
Рисунок 4. ПТМ тубулина в хоаноцитах интактной стенки тела. (a) ацетилированный тубулин в веретенах деления и пост-митотических хоаноцитах; (b) постепенное деацетилирование сети МТ в пост-митотических хоаноцитах. КЛСМ, максимальные проекции интенсивности. Anti-α-tubulin antibodies (фиолетовый), anti-acetylated-α-tubulin (жёлтый), DAPI (ядра, циановый). Масштабный отрезок: 5 мкм.
В статье также описана архитектура сети МТ в клетках мезохила. В склероцитах (клетках, синтезирующих спикулы) длинные переплетающиеся изогнутые пучки МТ направлены вдоль главной оси клетки, параллельно стресс-фибриллам отходят от единственного ЦОМТа вблизи ядра. В больших амебоидных клетках архитектура сети МТ также радиальная, а пучки МТ заходят в длинные ветвящиеся лопободиальные выросты этих клеток (Рис. 5). В склероцитах и больших амебоидных клетках не было обнаружено ацетилированных и детирозинированных МТ. Зато окрашивание на детирозинированный тубулин позволило авторам обнаружить новый тип клеток мезохила: маленькие округлые клетки с высоким ядерно-цитоплазматическим соотношением (Рис. 5). Их микротрубочки состоят только из детирозированного тубулина, полностью колокализованного с тотальным α-тубулином. В ядрах некоторых из них наблюдается конденсированный хроматин.
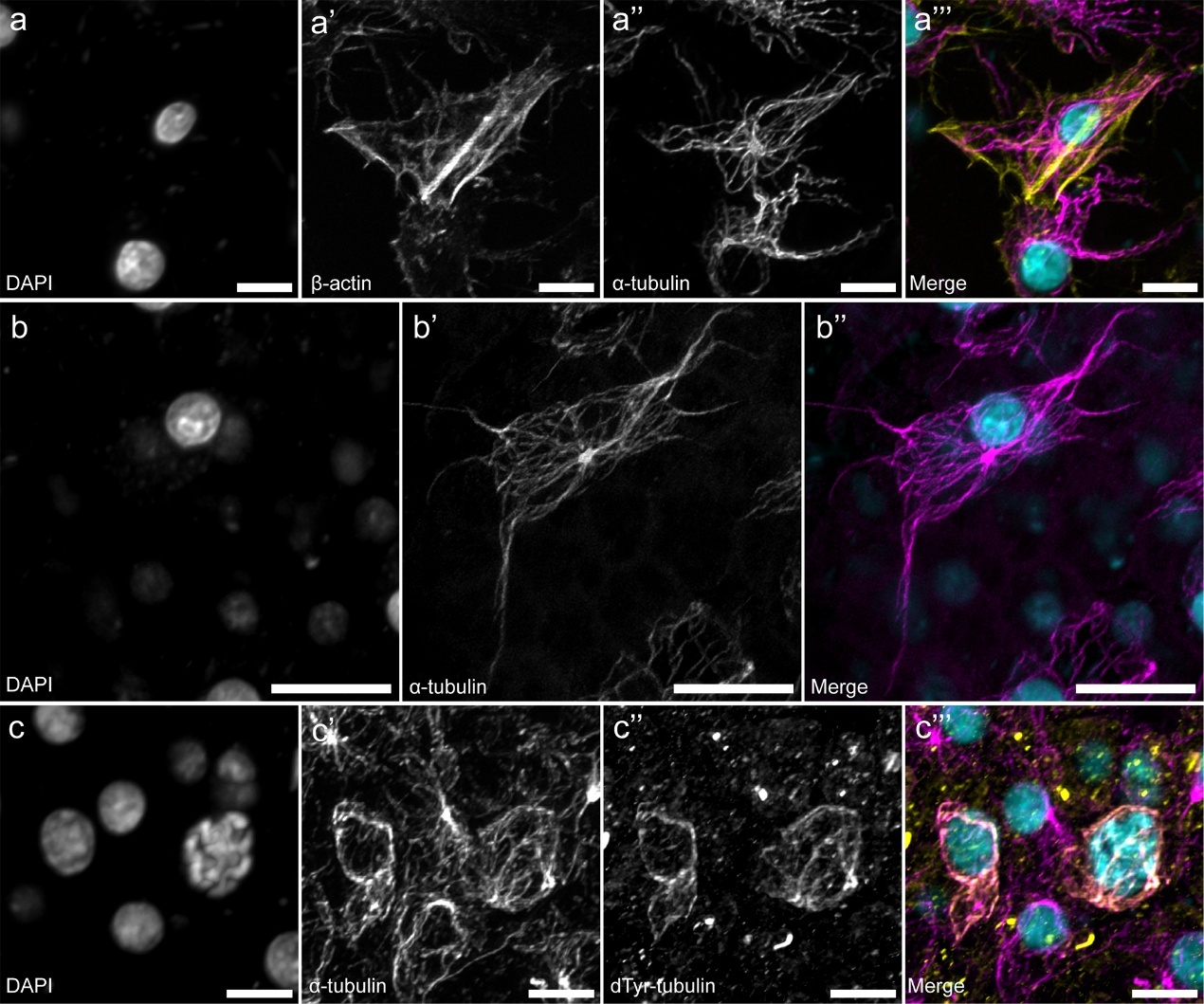
Рисунок 5. Микротрубочки и ПТМ тубулина в клетках мезохила. (a) Актиновые филаменты и МТ в склероците; (b) радиальная сеть МТ в большой амебоидной клетке; новая (обнаруженная авторами статьи) клетка мезохила, с полностью детирозинированными микротрубочками. КЛСМ, максимальные проекции интенсивности. Масштабный отрезок: (a, c) – 5 мкм; (b) – 10 мкм.
Перейдём к заключению. Что же новое и интересное для биологии развития и эволюционной биологии удалось обнаружить авторам статьи?
Описав архитектуру сети микротрубочек и ПТМ тубулина в этих сетях, авторы сопоставили эти данные с информацией по тубулиновому цитоскелету разнообразных беспозвоночных животных и клетками иммортализованных линий позвоночных. Они обращают внимание на то, что для пучков МТ в клетках губки характерна интересная особенность: они искривлены, сгибаются, могут менять свое направление. Известно, что сродство некоторых МТ-ассоциированных белков к таким микротрубочкам выше, что может обеспечивать локальное изменение их функций (Bechstedt et al., 2014). Обсуждая архитектуру сети МТ в хоаноцитах авторы подчеркивают универсальную для Metazoa апикобазальную полярность этих клеток, что подтверждает эпителиальную природу хоанодермы. Авторы также подчеркивают, что в морфогенезах, обеспечивающих регенерацию стенки тела губки, на клеточном уровне задействована система МТ. Известно, что стабилизация МТ способствует распластыванию клетки, регулирует транспорт внутри неё, обеспечивает поддержание старых и формирование новых межклеточных контактов. Кроме того, авторы отмечают, что динамичность сети микротрубочек в клетках губки может обеспечивать необходимую животному пластичность тканей, что способствует регенерации.
Авторы статьи также впервые проанализировали "тубулиновый код" губки и сделали несколько интересных выводов.
(1) Тирозинизированный тубулин является наиболее распространенной формой ПТМ, в то время как детирозинированные и ацетилированные МТ встречаются редко.
(2) Ацетилированный тубулин был обнаружен в веретене деления, аксонеме жгутика и сети МТ пост-митотических хоаноцитов. Известно, что ацетилирование обеспечивает стабильность МТ, а также связывание динеина, а значит эффективный транспорт везикул (Achler et al., 1989). Авторы предполагают, что ацетилированные МТ в пост-митотических хоаноцитах сохраняются, чтобы помочь дочерним клеткам установить контакт со своими соседями и завершить процесс поляризации. Кроме того, сохранение ацетилированных микротрубочек позволяет быстро сформировать жгутик и вернуться к работе в качестве клеток водоносной системы.
(3) Интерфазные МТ в клетках губок очень динамичны. В таких условиях ПТМ (являющиеся признаком “старых” микротрубочек) не могут накапливаться. Обнаруженные ПТМ связаны с определенными типами клеток, что может указывать на выполнение ими специфичных функций, зависящих от типа клетки.
(4) Морфогенезы, обеспечивающие регенерацию у известковых губок, зависят не только от реорганизации актиновых филаментов, но и от изменения архитектуры сети интерфазных МТ. МТ при регенерации обеспечивают стабилизацию направления миграции клеток, а также внутриклеточный транспорт, необходимый для установления полярности клеток и формирования межклеточных контактов.
Опубликованное исследование является еще одним шагом к пониманию сложных и эволюционно консервативных взаимодействий между элементами цитоскелета в клетках Metazoa, ответственных за подвижность клеток и изменение их формы.
- Achler, C., Filmer, D., Merte, C., & Drenckhahn, D. (1989). Role of microtubules in polarized delivery of apical membrane proteins to the brush border of the intestinal epithelium. The Journal of Cell Biology, 109(1), 179–189. https://doi.org/10.1083/jcb.109.1.179
- Bechstedt, S., Lu, K., & Brouhard, G. J. (2014). Doublecortin Recognizes the Longitudinal Curvature of the Microtubule End and Lattice. Current Biology, 24(20), 2366–2375. https://doi.org/10.1016/j.cub.2014.08.039
- Kirfel, G., & Stockem, W. (1997). Detection and cytoplasmic localization of two different microtubule motor proteins in basal epithelial cells of freshwater sponges. Protoplasma, 196(3–4), 167–180. https://doi.org/10.1007/BF01279565
- Lavrov, A. I., Bolshakov, F. V., Tokina, D. B., & Ereskovsky, A. V. (2018). Sewing up the wounds: The epithelial morphogenesis as a central mechanism of calcaronean sponge regeneration. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 330(6–7), 351–371. https://doi.org/10.1002/jez.b.22830
- Ludueńa, R. F., Shooter, E. M., & Wilson, L. (1977). Structure of the tubulin dimer. Journal of Biological Chemistry, 252(20), 7006–7014. https://doi.org/10.1016/S0021-9258(19)66927-9
- Mitchison, T., & Kirschner, M. (1984). Dynamic instability of microtubule growth. Nature, 312(5991), 237–242. https://doi.org/10.1038/312237a0
- Shida, T., Cueva, J. G., Xu, Z., Goodman, M. B., & Nachury, M. V. (2010). The major α-tubulin K40 acetyltransferase αTAT1 promotes rapid ciliogenesis and efficient mechanosensation. Proceedings of the National Academy of Sciences of the United States of America, 107(50), 21517–21522. https://doi.org/10.1073/pnas.1013728107
- Skorentseva, K. V, Bolshakov, F. V, Saidova, A. A., & Lavrov, A. I. (2023). Regeneration in calcareous sponge relies on ‘purse-string’ mechanism and the rearrangements of actin cytoskeleton. Cell and Tissue Research. https://doi.org/10.1007/s00441-023-03810-5
- Verhey, K. J., & Gaertig, J. (2007). The tubulin code. In Cell Cycle (Vol. 6, Issue 17, pp. 2152–2160). Taylor and Francis Inc. https://doi.org/10.4161/cc.6.17.4633
- Wachtmann, D., & Stockem, W. (1992). Significance of the cytoskeleton for cytoplasmic organization and cell organelle dynamics in epithelial cells of fresh-water sponges. Protoplasma, 169(3–4), 107–119. https://doi.org/10.1007/BF01323610
Новость подготовила
© Ксения Скоренцева, аспирант лаборатории эволюции морфогенезов.
18.04.2025
Работа была выполнена при поддержке гранта
РНФ №23-74-10005