 © Богуславский Дмитрий Викторович, старший научный сотрудник лаборатории нейробиологии развития ИБР РАН, кандидат биологических наук. Область научных интересов: молекулярная нейробиология, этология, апиология, социобиология.
© Богуславский Дмитрий Викторович, старший научный сотрудник лаборатории нейробиологии развития ИБР РАН, кандидат биологических наук. Область научных интересов: молекулярная нейробиология, этология, апиология, социобиология.Кросс-модальное различение — это распознавание, при котором объекты, представленные в одной модальности (например, зрительной), идентифицируются с использованием другой модальности (например, осязательной) через определенный временной промежуток. Кросс-модальное различение было показано через зрение и осязание у людей (1), человекообразных обезьян (2), обезьян (3) и крыс (4); через зрение и слух у дельфинов (5); через зрение и электрическую чувствительность у рыб (6). Способность распознавать объекты через разные модальности выгодно и адаптивно, позволяя полнее и лучше воспринимать окружающую среду в разных условиях. Кросс-модальное различение показывает, что животное имеет связанные «ментальные образы» объектов.
Группа из Великобритании под руководством известного немецкого нейроэтолога Ларса Читтки для поиска кросс-модального различения у беспозвоночных использовала очень удобный для этой цели объект – обыкновенного шмеля, который, как известно, не только хорошо ориентируется в светлое время суток благодаря паре сложных фасеточных глаз, но и легко ориентируется в темноте гнезда с помощью пары антенн (усиков, сяжек).
Авторы исследования обучили 44 шмеля находить корм (вода с 50% сахарозы) в темной комнате. Корм размещали в выемке верхней части либо шара, либо куба. Для отрицательного подкрепления использовался раствор хинина. В каждой чашке Петри было по четыре либо шара, либо куба (рис. 1А). Затем шмелей разделили на две группы. Первую группу (21 особь) использовали для того, чтобы проверить, могут ли шмели распознать предмет на ощупь в темноте: насекомые значительно (p=4,5×10-15) больше времени проводили у тех фигур, в которых до этого находили угощение, даже в том случае, если в них ничего не было, ориентируясь, по-видимому, только на их форму (рис. 1С). Вторую группу из 23 шмелей пустили в уже освещенную комнату с теми же фигурами, только прикрытыми прозрачным стеклом со всех сторон: так, чтобы шмели могли их видеть, но не могли трогать (рис. 1В). В этом случае насекомые также значительно (p=9,8×10-16) больше времени проводили у тех блоков, в которых ранее находили угощение — даже с учетом того, что не могли их потрогать, а видели их впервые (рис. 1D).
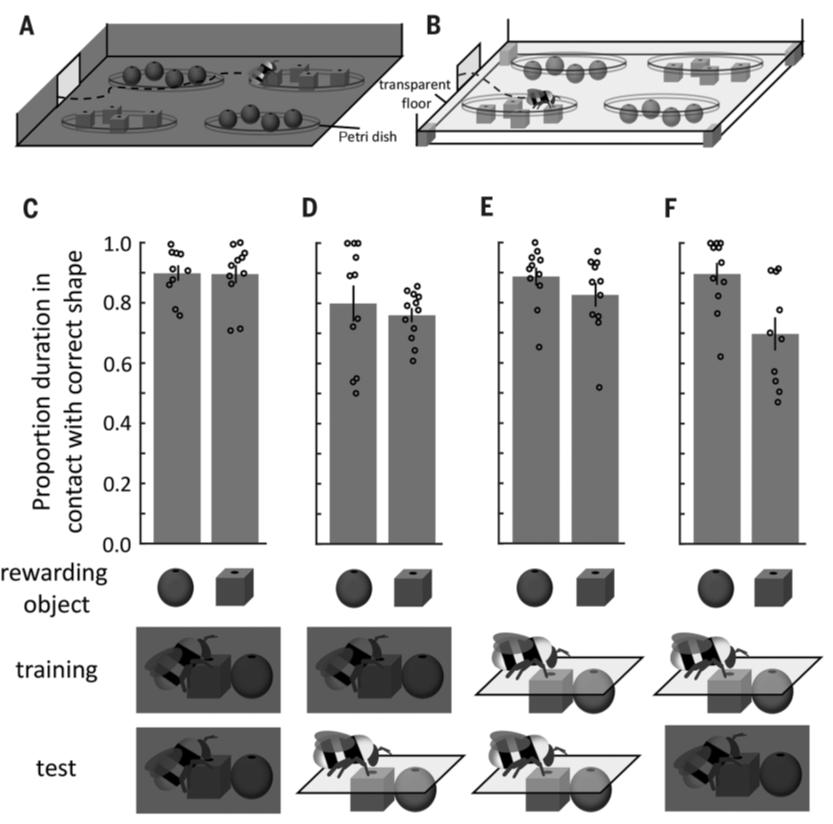
Рис 1.
Во второй части исследования учеными было проведено обратное кросс-модальное различение — визуально-тактильное. Было обучено 43 шмеля искать корм в отверстиях стекла над фигурами в светлой комнате с помощью хоботка. При этом насекомые не могли прикасаться к самим фигурам своими антеннами. Затем 22 шмеля были протестированы на тех же самых кормушках, но без корма (рис. 1Е). Эти шмели значительно (p=1,36×10-23) больше времени проводили у кормушек над фигурами, у которых до этого получали угощение. Остальных шмелей затем протестировали в темной комнате, где нужно было ориентироваться с помощью осязания. Шмели значительно (p=0,0056) больше времени проводили у фигур, из которых ранее получали корм (рис. 1F).
Эти результаты показали, что после обучения различать объекты, используя только одну сенсорную модальность, шмели могут различать эти же объекты с помощью другой сенсорной модальности. При этом шмели не распознают объекты, если они их не видели или не могли ощупать (рис.2). Была проверена способность шмелей различать два объекта в темноте, не имея возможности их коснуться. Одна группа насекомых (n = 20) была обучена различать фигуры визуально в освещенной арене (рис. 1В), а другая группа (n = 21) обучена различать фигуры тактильно в темной арене (рис. 1А). Впоследствии во время теста в темноте и без возможности касаться фигур (рис. 2А), обе группы не показали разницы в продолжительности нахождения над фигурами (рис. 2, В и С).
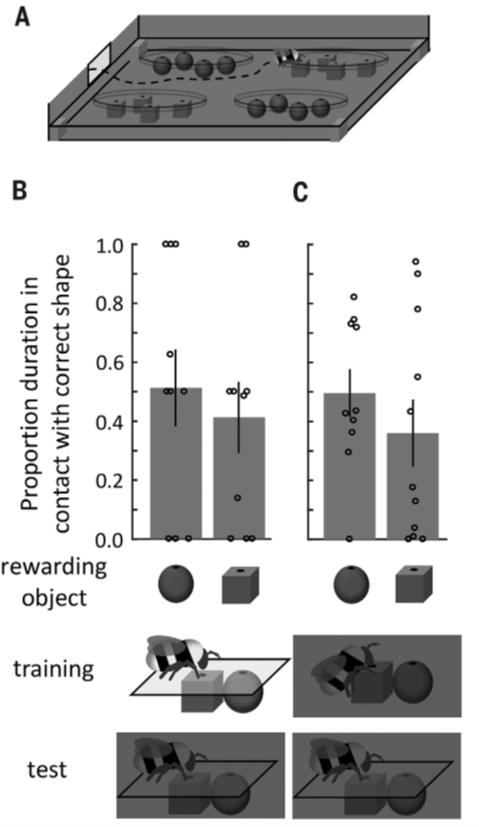
Рис 2.
Восприятие объектов является результатом интеграции информации от различных сенсорных каналов (6, 11). Два центра высокого уровня в протоцеребруме насекомых, грибовидные тела и центральный комплекс, получают как зрительную, так и сенсорную информацию (12-14). Авторы публикации предполагают, что эти области – возможные кандидаты на сенсорную интеграцию кросс-модального распознавания объектов. Кросс-модальное распознавание требует выполнения нескольких условий: два разных сенсорных канала должны получать информацию об одном и том же свойстве объекта (например, формы); различная по времени и способу кодирования информация должна быть идентифицирована как связанная с одним и тем же свойством объекта; информация должна хранится в нейронном представительстве, доступном обоим сенсорным каналам (8). Решают ли шмели задачу, имея представление о целой форме объекта (куб или сфера) или о локальных свойствах объекта (изогнутый или плоский край) остается пока неизвестным. В любом случае, эти эксперименты показывают, что шмели способны распознавать свойства объектов, получая различную по времени и способу кодирования информацию. Насекомые демонстрируют своего рода информационную интеграцию, что требует модально-независимого внутреннего представительства (7, 9, 10). Всё это говорит о том, что похожее на людей и других животных с крупным мозгом, поведение насекомых связано с интеграцией информации из нескольких сенсорных каналов в целостный образ восприятия окружающего их мира (6, 15, 16).
1. Gaydos, H. Intersensory transfer in the discrimination for form. Amer. J. Psychol., 1956, 69, 107–110.
2. Davenport, R. K., and Rogers, C. M. Intermodal equivalence of stimuli in apes. Science, 168, 279–280, 1970.
3. Cowey A, Weiskrantz L. (1975) Demonstration of cross-modal matching in rhesus monkeys, Macaca mulatta. Neuropsychologia 13:117–120.
4. Winters, B.D. & Reid, J.M. A distributed cortical representation underlies crossmodal object recognition in rats. Journal of Neuroscience, 30, 6253-6261. (2010).
5. Herman LM, Pack AA, Hoffmann-Kuhnt M. Seeing through sound: dolphins (Tursiops truncatus) perceive the spatial structure of objects through echolocation. J Comp Psychol 112:292–305. (1998).
6. C. Spence, in Stevens’ Handbook of Experimental Psychology and Cognitive Neuroscience (American Cancer Society, 2018), pp. 1–56.
7. L. M. Herman, A. A. Pack, M. Hoffmann-Kuhnt, Seeing through sound: Dolphins (Tursiops truncatus) perceive the spatial structure of objects through echolocation.J. Comp. Psychol. 112, 292–305 (1998).
8. S. Schumacher, T. Burt de Perera, J. Thenert, G. von der Emde. Cross-modal object recognition and dynamic weighting of sensory inputs in a fish.Proc. Natl. Acad. Sci. U.S.A. 113, 7638–7643 (2016).
9. B. E. Stein, M. A. Meredith. Multisensory integration. Neural and behavioral solutions for dealing with stimuli from different sensory modalities. Ann. N. Y. Acad. Sci. 608, 51–70 (1990).
10. Kosslyn, S. M. (1995). Mental imagery. In S. M. Kosslyn & D. N. Osherson (Eds.), An invitation to cognitive science. Visual cognition: An invitation to cognitive science (p. 267–296). The MIT Press.
11. B. De Gelder, P. Bertelson. Multisensory integration, perception and ecological validity. Trends Cogn. Sci. 7, 460–467 (2003).
12. P. G. Mobbs, The connections and spatial organization of the mushroom bodies. Philos. Trans. R. Soc. London B Biol. Sci. 298, 309–354 (1982).
13. N. J. Strausfeld. J. Comp. Neurol. 450, 4–33 (2002).
14. K. Pfeiffer, U. Homberg, Organization and functional roles of the central complex in the insect brain. Annu. Rev. Entomol. 59, 165–184 (2014).
15. L. Mudrik, N. Faivre, C. Koch, Trends Cogn. Sci. 18, 488–496 (2014).
16. O. Deroy et al., Multisens. Res. 29, 585–606 (2016).
Новость подготовил © Богуславский Д.В.
18.03.2020