
Область научных интересов: нейроэтология, нейробиология, нейротрансмиттеры, поведение.

Область научных интересов: нейробиология, биология поведения.
Когнитивные функции можно тренировать, также как и двигательные, это известно давно. Но вот о том, какой именно след остается в обученной нервной системе, обеспечивающий ее более высокую пластичность и способность учиться в будущем известно крайне мало. В очередной раз моллюск Большой прудовик зарекомендовал себя как объект, на котором можно решать самые удивительные задачи, в том числе и эту. Группа Джорджа Кеменеша (Великобритания) уже больше десяти лет исследует клеточные и молекулярные механизмы ассоциативного обучения у этого вида. Ими создана модель, в которой ассоциативная связь между новым нейтральным веществом и пищевым подкреплением устанавливается сразу, после единственного сочетания, и надолго. Последняя их работа свидетельствует о том, что новая ассоциация формируется легче у улиток, которые недавно находились в процессе другого обучения (Crossley et al., 2023).
Авторы работали на конкретной, изученной ранее нервной сети пищевого поведения (пищевой генератор). В их экспериментах происходит выработка ассоциативной положительной связи между нейтральными веществами (амилацетат или гамма-ноналактон) и пищевым подкреплением (0.33 % сахароза). Как отмечено выше, достаточно одного такого сочетания, чтобы улитка в последствии реагировала на нейтральный фактор движением радулы (пищевой аппарат прудовика), то есть устанавливала ассоциативную связь, обучалась. Если использовать сахарозу в три раза более слабой концентрации (0.11%), обучение не происходит. Сила подкрепления имеет значение. Но если сначала обучить улитку с сильным подкреплением, то затем та же особь уже способна сформировать условный рефлекс (ассоциацию) и с другим нейтральным стимулом, подкрепленным слабой концентрацией сахарозы.
Основная задача при таком подходе – очень аккуратно провести контрольные эксперименты, показывая, что «слабое» обучение возможно тогда и только тогда, когда ему предшествовало «сильное». Авторам удалось доказать, что (1) такое «слабое» обучение не зависит от того, какое из двух нейтральных веществ использовалось первым, т.е. в процедуре с сильным подкреплением; (2) что обучение с сильным подкрепление не вызывает неспецифической положительной реакции на другое нейтральное вещество, то есть слабое подкрепление необходимо, чтобы сформировалась реакция на второе нейтральное вещество (3) что обучение с сильным подкреплением нельзя заменить на одно подкрепление, то есть высокая концентрация глюкозы перед сочетанием нейтрального стимула со слабым подкреплением не приводит к формированию ассоциативной связи. Зато (4) 0.11% сахарозу можно заменить на 0.11% L-серин, другое слабое пищевое подкрепление (Рис. 1)! Последнему результату авторы придают большое значение, ведь он показывает, что не только безусловный стимул, но и подкрепление может быть другим в слабой серии, а обучение все-равно происходит. Значит, мы действительно, имеем дело с повышением некоторой общей предрасположенности к установлению новых связей!
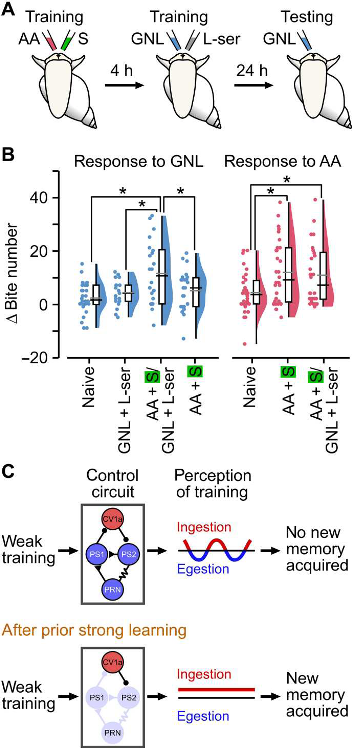
Рис. 1. (A) Схема двойной тренировки с использованием отдельных условного и безусловного (CS + US) стимулов для каждой тренировки. Сильная тренировка заключалась в сочетании
амилацетата (AA) в качестве условного стимула (CS) с сахарозой (S, 0,33%), которая была в
качестве безусловного стимула (US). Четыре часа спустя животные получали слабую тренировку
гамма-нолалактоном (GNL) в виде CS в паре с L-серином (0,11%) в качестве US. Через день
животных проверяли на реакцию CS.
(B) Сильная тренировка за 4 часа до слабой тренировки с L-серином в US привела к значительному
усилению реакции на GNL по сравнению с наивным, только слабым или только сильным
(наивные, n = 26; только слабые, n = 19; сильный за 4 часа до слабого, n = 22; только сильные, n =
24). Сильная тренировка, сопровождаемая слабой тренировкой, также привела к значительному
усилению реакции на АА по сравнению с наивным, но не только сильным (наивный, n = 32; только
сильные, n = 30; сильный за 4 часа до слабого, n = 26).
(C) Схематическое изложение предлагаемого механизма, т.е. как вызванный обучением сдвиг в
состоянии сети может изменить восприимчивость к последующему слабому обучению и улучшить
приобретение новой памяти. Слабое обучение само по себе воспринимается как
неоднозначное/нестабильное из-за конкурентных взаимодействий между интернейронами,
управляющими заглатыванием пищи и выплевыванием, что приводит к переходу животного
между двумя видами поведения без какого-либо приобретения памяти. Однако предшествующая
сильная тренировка стабилизирует состояние сети и смещает активность в сторону схемы
поглощения, изменяя восприятие последующей слабой тренировки и улучшая приобретение
новых воспоминаний. h - часы.
В чем может быть биологический смысл? Авторы предлагают объяснение удивительно сходное с нашей интерпретацией ранее полученных результатов о сохранении нейронального следа повышенной моторной активности у этого вида (Dyakonova et al., 2019). Они предполагают, что появление нового сигнала, связанного с биологически важным, означает для организма меняющиеся условия, в которых можно ожидать появление и других новых факторов, требующих научения. По их выражению однократное обучение становится сигналом о наступлении целого периода «богатого на обучение». Довольно серьезная заявка, если учесть объект, на котором получены результаты. В любом случае, результаты стали очередным свидетельством в пользу существования механизма формирования предсказаний, причем не только конкретных, но и обобщенных!! у относительно просто устроенного организма.
Далее авторы, пользуясь преимуществами выбранного объекта, у которого хорошо известны нейроны, формирующие пищевое поведение, провели электрофизиологические исследования, чтобы выявить механизм реализации такого комбинированного формирования памяти на клеточном уровне. Они сосредоточились на тех нейронах пищевого поведения, которые запускают фиктивный цикл пищевого ритма в буккальных ганглиях улитки, а также на тех элементах, которые меняют этот пищевой цикл так, что улитка вместо заглатывания пищи начинает отрыгивать пищу наружу. Такая работа пищевого генератора позволяла оценить, насколько та или иная группа особей предрасположена к поеданию или отказу от пищи, и насколько поменялись предпочтения у экспериментальной группы по сравнению с контрольной. С помощью этих экспериментов были выявлены изменения в нескольких нейронах, известных ранее, как влияющих на запуск пищевого поведения, а также были выявлены новые нейроны, участвующие в механизме, который лежит в основе формирования «слабого» обучения в случае, если перед ним было «сильное» обучение.
Далее авторы использовали знание эргичности нейронов, участвующих в пищевом поведении. Они блокировали сульпиридом действие дофаминэргических нейронов, один из которых прямо участвовал в управлении пищевого поведения. Сульпирид блокировал торможение основного генератора пищевого ритма, т.е. один из идентифицированных дофаминовых нейронов переставал подавлять другой нейрон, управляющий активацией генератора. Авторы выяснили, что сульпирид действует на активность пищевого ритма не только непосредственно.
В схеме двойного, «сильного» и следующего за ним «слабого», обучения, сульпирид мог заменять «сильное» условное обучение на клеточном уровне. После него «слабое» обучение так же оставляло памятный след, т.е. формировалась долгосрочная память. Таким образом, отключение вытормаживающей связи в управлении пищевым генератором так же способствовало тому, что «слабое» обучение улитки пищевому стимулу не проходило для неё бесследно, в отличие от того, как это происходило в случае одного только «слабого» обучения.
Вместе с тем, сульпирид не добавлял каких-либо изменений в случаях только «сильного» обучения. Несмотря на то, что «сильное» обучение приводит к изменениям в тех же нейронах и связях пищевого генератора, сульпирид не влиял на эффективность формирования памятного следа при «сильном» обучении. Это дало авторам основание сделать вывод, что хотя все события, которые они рассматривают, происходят на одних и тех же нейронах генератора пищевого паттерна, можно полагать, что в этой схеме идут два процесса. Первый - собственно сам процесс изменения сетей генерации паттерна. Эти изменения происходят в момент химических и электрических воздействий на элементы управления пищевого генератора и сохраняются как памятные следы в изменениях состояния этих элементов. Второй – собственно изменение восприимчивости к последующему обучению, которое непрямо активируется в момент «сильного» обучения. Описанные два процесса, по логике авторов, реализуются независимо, а в случае, когда обучение представляет собой ряд независимых актов, эти два процесса развиваются одновременно. Авторы полагают, что восприимчивость к обучению – это отдельное свойство, которое проявляется в представленных экспериментах с улиткой в том, что улитка, находясь в среде с обогащенной обучающими событиями средой (в нашем случае – с множеством пищевых стимулов), становится более восприимчивой к обучению. И это не зависит, по большому счету, от того, какие конкретно эти стимулы и насколько они друг с другом связаны.
Таким образом, авторы на элементарном уровне системы (на уровне отдельных нейронов) не только доказали наличие связи между двумя последующими обучениями, но и выявили первые механизмы этой связи.
Новость подготовили
© В.Е.Дьяконова
© И.А.Чистопольский
15.11.2023