 © Дьяконова Варвара Евгеньевна
доктор биологических наук, ведущий научный сотрудник,
лаборатория нейробиологии развития. Область научных интересов:
нейроэтология, нейробиология, нейротрансмиттеры, поведение.
© Дьяконова Варвара Евгеньевна
доктор биологических наук, ведущий научный сотрудник,
лаборатория нейробиологии развития. Область научных интересов:
нейроэтология, нейробиология, нейротрансмиттеры, поведение.
Развитие клеточных технологий уже позволяет получать ткане-специфические клетки из стволовых эмбриональных или дедиферинцированных взрослых клеток. В отношении нервной ткани, этот шаг был наиболее труден, поскольку функциональным может быть только нейрон с конкретной трансмиттерной специфичностью, набором рецепторов и электрическими свойствами. Нервная система неоднородна (гетерогенна), включает большое число фенотипически различных нейронов, и эта неоднородность - ее важнейшее функциональное свойство. Базовым функциональным элементом нервной системы можно рассматривать гетерохимический эндогенно активный нейронный ансамбль.
Многие формы поведения обеспечиваются такими нейрональными ансамблями, способными к переформатированию выходного паттерна. К их числу относятся центральные генераторы паттерна (ЦГП) локомоции большинства животных. Понятие нейрональный ансамбль занимает все более уверенную позицию не только в нейробиологии моторных программ, но и в механизмах восприятия, памяти и обучения. Уже давно было известно, что нейроны, формирующие такие ансамбли, принадлежат к разным химическим и электрическим фенотипам. В естественных условиях формирование фенотипических свойств нейрона в значительной степени определяется актуальной гетерохимической межклеточной средой (нишей), в которой оказываются клетки-предшественники. Эта среда, в свою очередь, является результатом секреции разных сигнальных молекул уже существующими клеточными структурами.
Значимость фенотипических различий нейронов для формирования и перестройки ритмической активности ансамбля остается не вполне понятой на теоретическом уровне: не существует моделей, которые бы умели предсказывать характер выходной активности в зависимости от качественного и количественного состава ансамбля.
В 2017 году была предпринята попытка подойти к решению этой задачи экспериментальным путем. Штернфельд и коллеги (Sternfeld et al. 2017) разработали новый метод, который позволяет получать из эмбриональных стволовых клеток фенотипы нейронов, входящие в состав спинального ЦГП локомоции позвоночных, и формировать из таких нейронов искусственные ансамбли с разным качественным и количественным составом (Рис.1, 2). Эти ансамбли назвали «циркутоиды» от английского circuit (контур, цикл, нечто, способное к генерации повторяющегося ритмичного паттерна активности).
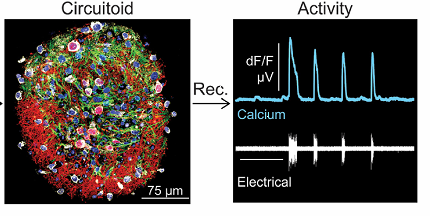
Рис. 1. Циркутоид, регистрация изменений концентрации кальция и электрической активности в нем.
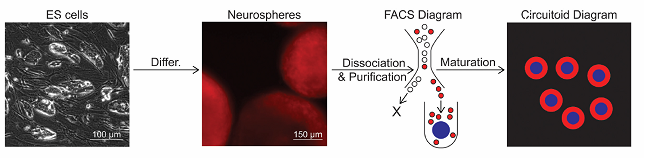
Рис. 2. Получение циркутоида. Получение нейросфер из эмбриональных стволовых клеток, их дальнейшая диссоциация и очистка, отбор нейронов нужных фенотипов, подсадка центрообразующей глиальной клетки (синий круг), вокруг которой происходит рост циркутоида.
Было использовано 4 основных нейрональных фенотипа: глутаматергические интернейроны двух несколько отличающихся по электрогенезу типов, гамк/глицинергические «тормозные» интернейроны, а также мотонейроны, синтезирующие ацетилхолин. Общее количество нейронов в циркутоиде варьировало от 5000 до 100000 клеток.
Основные результаты этой работы заключаются в следующем.
- Показано, что циркутоиды, состоящие из четырех основных фенотипов нейронов, способны к генерации эндогенной ритмической активности.
- Эта активность модулируется нейротрансмиттерами, которые активируют локомоторный ритм в естественных условиях, например, серотонином. При этом частота ритма циркутоида оказалась близкой к частоте естественного локомоторного ЦГП.
- К генерации эндогенного ритма также оказались способны циркутоиды, состоящие более чем на 98 % из глутаматергических интернейронов. При этом не было различий между ритмами, генерируемыми циркутоидами, отличающимися на порядок по количеству нейронов.
- Ритм генерировался и циркутоидами, состоящими из мотонейронов, однако более медленный и менее стабильный.
- Циркутоиды, состоящие из ГАМК/глицинэргических интернейронов были не способны к генерации собственного ритма.
- Авторы также исследовали вклад тормозных интернейронов в электрогенез циркутоида, варьируя соотношение ритм-генерирующих и тормозных нейронов в циркутоиде (Рис. 3). Повышение количества тормозных интернейронов в циркутоиде с глутаматергическими интернейронами приводило к увеличению частоты генерации эндогенного ритма, эффект можно было снять добавлением антагонистов ГАМК (Рис. 3D).
- Иной эффект наблюдался при добавлении тормозных интернейронов в циркутоиды с ацетилхолиновыми мотонейронами. В этом случае наблюдалось разделение единого ансамбля на подансамбли, генерирующие свои ритмы (Рис. 3Е).
- Наконец, добавление глуматных интернейронов в ацетилхолин/гамкергические циркутоиды приводило к восстановлению единой сети генерации ритма с частотой, характерной для глумат- гамкергических циркутоидов (Рис. 3F)
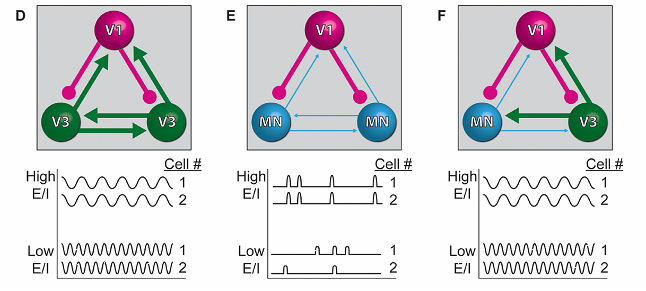
Рис. 3. Схематическое изображение зависимости ритмической активности от фенотипического состава циркутоида и предположительное объяснение полученных эффектов. Внесение тормозных нейронов в циркутоиды, состоящие из возбуждающих нейронов V3, приводит к учащению ритма, а в циркутоиды, состоящие из мотонейронов с более слабыми возбуждающими связями, к распаду ритма циркутоида на отдельные ритм-генерирующие ансамбли. Более сильные возбуждающие нейроны V3, добавленные в последние циркутоиды, переподчиняют циркутоид единому ритму.
С моей точки зрения, эта работа указывает на существующий теоретический пробел в нейробиологии: если бы мы, действительно, понимали, каким образом нейроны объединяются в ансамбли, и как формируется выходной паттерн ансамбля, все эффекты, описанные на уровне циркутоида, могли быть предсказаны математической моделью, которая бы учитывала функционально-значимые характеристики клеток. Еще более обидно, что отсутствие такой теоретической базы не позволяет в полной мере использовать возможности этого нового метода. Он был бы идеален для экспериментальной проверки конкретных гипотез и адекватности математических моделей нейрональных ансамблей. Без этого полученные результаты, хотя и представляют несомненный интерес, все же остаются в рамках эмпирической и феноменологической биологии. Клеточные технологии в данном случае опережают теоретическую фундаментальную нейробиологию, которая пока оказалась не в состоянии использовать их методический ресурс.
Новость подготовила © 2017 Дьяконова В.Е.
15.06.2017