 © Карпова Ярослава Дмитриевна
к.б.н., научн. сотр. лаборатории биохимии процессов онтогенеза.
Область научных интересов: биология развития, протеасомная биология, иммунологическая толерантность.
© Карпова Ярослава Дмитриевна
к.б.н., научн. сотр. лаборатории биохимии процессов онтогенеза.
Область научных интересов: биология развития, протеасомная биология, иммунологическая толерантность.
В последнем номере Science сразу две статьи посвящены роли протеасом в нахождении пары, образовании синапса и гомологичной рекомбинации хромосом в мейозе. До настоящего времени некоторые работы указывали на возможное участие протеасом в этих процессах, однако ранее их роль не была четко определена.
При подготовке к первому делению мейоза хромосомы объединяются в соответствующие гомологичные пары, в цепи ДНК делаются двунетевые разрезы, которые являются стартовой точкой образования гомологичных пар, хиазм и кроссинговера. Хромосомы имеют множество потенциальных стартовых сайтов, но лишь некоторые из них в действительности становятся началом кроссинговера. Prasada Rao с соавторами обнаружили, что центральную роль в выборе таких сайтов играют белки SUMO, убиквитин и протеасомы.
Известно, что навешивание специальными лигазами небольшого белка SUMO на белок-реципиент способствует его стабилизации и маркирует правильную внутриклеточную локализацию. Наоборот, навешивание похожего на SUMO белка убиквитина (точнее цепочки из молекул убиквитина) является сигналом на уничтожение в протеасоме. Координационной работе двух систем авторы и приписывают ключевую роль в регуляции профазы первого мейоза и успешного проведения кроссинговера. Используя методы иммунофлуоресцентного окрашивания ингибиторов систем SUMOляции, убиквитинирования и протеасомного гидолиза, они изучали сперматогенные клетки семенников нормальных и нокаутных мышей.
На основе полученных результатов авторы предложили следующую схему происходящих событий. Когда хромосомы образуют пару, HEI10 SUMO-лигаза метит регуляторы рекомбинации в области двунетевых разрывов ДНК. Если заблокировать этот процесс, то ни один потенциальный сайт не участвует в кроссинговере и мейоз останавливается. Далее, в некоторых сайтах SUMO заменяется на убиквитин с помощью RNF212-лигазы и, меченный таким образом, фактор деградируется протеасомой, двойной разрыв репарируется и кроссинговер не идет. Важно, что убиквитинирование без предшествующей SUMOляции не происходит. Таким образом, именно баланс этих двух систем приводит к нужному количеству точек кроссинговера, минимум одному на хромосому.
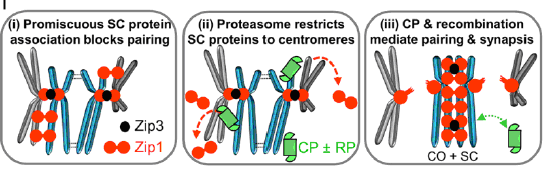
Во второй статье Ahuja с соавторами исследовали участие протеасом в первом делении мейоза дрожжей. В своей работе они пришли к похожим выводам, показали участие протеасом в процессе гомологичной рекомбинации и инициации кроссинговера в местах двойных разрывов ДНК и даже выявили связующую роль именно ZIP3-лигазы (гомолога человеческой RNF-212-лигазы).
Кроме этого, еще одним важным выводом из их работы стало определение роли протеасом в нахождении хромосомами своей гомологичной пары. При ингибировании гидролиза в протеасоме у дрожжей в норме и у нокаутов по одной из субъединиц протеасом значительно возрастает количество центромерно-спаренных негомологичных хромосом. По-видимому, перед образованием правильных пар хромосом происходит беспорядочное взаимодействие синаптических белков с хромосомами, которое ведет к временной стабилизации негомологичных пар. Протеасомы «снимают» их со всех неправильных мест, за исключением центромерных областей. Если плечи хромосом нерекомбинируют друг с другом, то контакт в центромерной области становится слабым и полностью дестабилизируется протеасомной деградацией связывающих белков. Если заблокировать это функционирование протеасом, то, во-первых, дальнейшее развитие идет по некроссоверному пути, во-вторых, хромосомы спариваются неправильно.
Выводы, полученные в обеих работах, не только имеют фундаментальную ценность, проясняя происходящие события на ключевых этапах мейоза, но и могут помочь в понимании причин нарушений образования половых клеток, вызывающих бесплодие и генетические патологии плода и у людей, поскольку происходящие события эволюционно достаточно консервативны.
 |
Новость подготовила © 13.02.2017 Карпова Я.