Vasileios Papdogiannis, Hugo J. Parker, Alessandro Pennati, Cedric Patthey, Marianne E. Bronner and Sebastian M. Shimeld
 © Гасанов Евгений Валерьевич,
ведущий научный сотрудник лаборатории нейробиологии развития. Кандидат химических наук.
© Гасанов Евгений Валерьевич,
ведущий научный сотрудник лаборатории нейробиологии развития. Кандидат химических наук.Интересы: биология развития, эволюционная биология, механизмы формирования нервной системы, молекулярная генетика.
Чувствительные ганглии головы, а именно чувствительные ганглии тройничного, вестибулярно-слухового (преддверно-улиткового) и эпибранхиального (языкоглоточного) черепно-мозговых нервов позвоночных представляют собой скопления сенсорных нейронов, передающих информацию непосредственно или от чувствительных клеток в головной мозг. Их формирование в эмбриогенезе происходит из черепных плакод, отделяющихся от эктодермы (чувствительные нейроны), и мигрирующих клеток нервного гребня головы (глия и некоторые нейроны тройничного ганглия). Их появление у позвоночных связано с выделением, собственно, головы на переднем конце тела предков позвоночных и объясняют их переходом от фильтрации к хищничеству (Lara-Ramirez et al., 2017). Логично, что ни самих чувствительных ганглиев головы, ни формирующих их черепных плакод и нервного гребня у ближайших хордовых родственников позвоночных, близких к их предковой форме - ланцетнков и оболочников - нет. Однако в обозреваемой работе (Papdogiannis et al., 2022) авторам удалось обозначить у низших хордовых элементы нервной системы, вероятно, гомологичные таковым, давшим начало чувствительным ганглиям головы у предков позвоночных.
Прежде всего авторы обозреваемой работы сконцентрировались на генах семейства Hmx, кодирующих гомеодомен-содержащие факторы транскрипции. Челюстноротые имеют четыре гена семейства: Hmx1, Hmx2, Hmx3 и SOHo, экспрессирующиеся в центральной нервной системе (ЦНС) и черепных плакодах, а также чувствительных ганглиях головы как их производных. При этом Hmx3 может служить маркером происходящих из плакод сенсорных нейронов чувствительных ганглиев головы. Было выявлено, что миноги - наиболее базальные, бесчелюстные позвоночные - имеют три гена семейства: HmxA, HmxB и HmxC, экспрессирующиеся в центральной, но не периферической нервной системе и производных черепных плакод, включая чувствительные ганглии головы. Видимо, такой тип экспрессии генов Hmx отражает предковое состояние для всех позвоночных.
Анализ экспрессии генов Hmx у ланцетника показал, что единственный ген также экспрессируется в ЦНС, но не в периферической нервной системе. Однако у оболочника Ciona intestinalis также единственный ген Hmx экспрессируется в субпопуляции периферических хвостовых биполярных нейронов, закладывающихся латеральнее нервной пластинки, что, как полагали ранее, роднило их с нервным гребнем (Smith et al., 2018).
Сравнение Hmx генов бесчелюстных (миноги и миксины) с позвоночными показало, что единственный Hmx ген хордовых удвоился у предка позвоночных, сформировав кластер из тандемно повторяющихся Hmx генов. Следующее удвоение одного из Hmx генов у бесчелюстных привело к формированию кластера из трёх генов: HmxA, HmxB и HmxC, тогда как у челюстноротых в результате полногеномного удвоения удвоился весь изначальный кластер из двух Hmx генов, сформировав два кластера по два гена семейства.
Кластер Hmx3-Hmx2 челюстноротых содержит два консервативных некодирующих элемента (CNE), чьи гомологи найдены и у бесчелюстных, что говорит об их возникновении при первоначальном удвоении единственного гена Hmx хордовых. Авторы протестировали экспрессию репортерной конструкции под контролем обоих CNE в трансгенной линии миног (рис.1). В обоих случаях CNE стимулировали экспрессию зелёного флуоресцентного белка (GFP) по типу Hmx генов, включая чувствительные ганглии головы. Авторы логично предполагают, что предковый единственный CNE, регулировавший первоначальный Hmx ген, в таком случае, также направлял его экспрессию по схожему паттерну. Однако протяжённый CNE оболочников стимулирует экспрессию Hmx как в центральной, так и периферической нервной системе и отличен по последовательности от CNE позвоночных. Однако CNE миноги, введённый в зиготу оболочника Ciona intestinalis с репортерной конструкцией (lacZ), показал экспрессию, совпадающую с таковой Hmx гена оболочника, включая ЦНС и субпопуляцию периферических хвостовых биполярных нейронов (рис. 1).
Предполагалось, что прилегающая к нервной пластинке эктодерма оболочников, дающая начало нервным клеткам, делится в передне-заднем направлении на закладки прото-черепных плакод и прото-нервного гребня позвоночных, соответственно (Horie et al., 2018). Однако экспрессия Hmx гена и репортерной конструкции под контролем CNE Hmx генов позвоночных хвостовыми чувствительными нейронами оболочника, формирующимися вдоль задней части нервной пластинки, говорит об их сходстве с чувствительными нейронами черепных нервов и, следовательно, гомологии этой области плакодам позвоночных. Поскольку головная эктодерма, прилегающая к нервной пластинке оболочников, уже была определена как гомолог обонятельных плакод позвоночных (Horie et al., 2018), возникает резонный вопрос о наличии у оболочников гомологов нервного гребня. Возможно, их отсутствие у оболочников связано со вторичным упрощением, которому подверглась группа в связи с неподвижным образом жизни. В этом смысле интересно выявить маркеры нервного гребня у ланцетника, как подвижного и менее специализированного представителя низших хордовых. Что же касается чувствительных ганглиев черепно-мозговых нервов позвоночных, то их гомологи присутствуют у самых примитивных хордовых, подразумевая эволюционную древность как их возникновения, так и генетической регуляции их формирования.
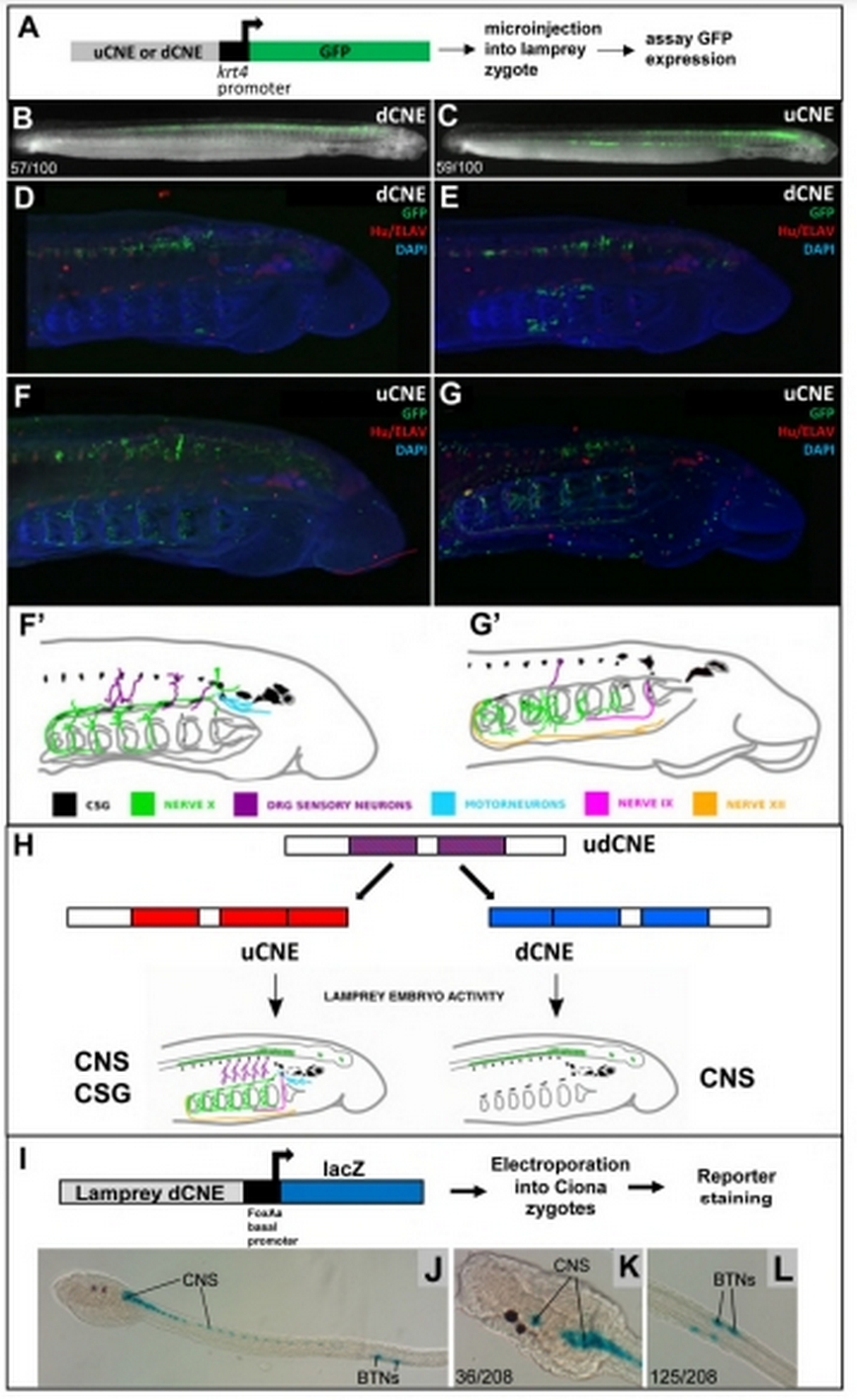
Рис. 1.
Активность CNE Hmx генов миноги в развивающихся миногах Petromyzon marinus (A-H) и оболочнике Ciona intestinalis (I-L). A, I - схема эксперимента, B, D, E и C, F, G - экспрессия GFP под контролем двух разных CNE миноги. H - схема эволюции двух CNE у позвоночных. CNS - ЦНС, CSG - чувствительные ганглии головы, BTNs - биполярные хвостовые нейроны оболочника.
1) Papadogiannis V, Pennati A, Parker HJ, Rothbächer U, Patthey C, Bronner ME, Shimeld SM. Hmx gene conservation identifies the origin of vertebrate cranial ganglia. Nature. 2022; 605(7911): 701-705. doi: 10.1038/s41586-022-04742-w.
2) Lara-Ramirez, R., Poncelet, G., Patthey, C. & Shimeld, S. M. The structure, splicing, synteny and expression of lamprey COE genes and the evolution of the COE gene family in chordates. Dev Genes Evol 227, 319-338, doi:10.1007/s00427-017-0591-6 (2017).
3) Smith, J. J. et al. The sea lamprey germline genome provides insights into programmed genome rearrangement and vertebrate evolution. Nat Genet 50, 270-277,
doi:10.1038/s41588-017-0036-1 (2018).
4) Horie, R. et al. Shared evolutionary origin of vertebrate neural crest and cranial placodes. Nature 560, 228-232, doi:10.1038/s41586-018-0385-7 (2018).
Новость подготовил
© Гасанов Е.
11.07.2022