
Научные интересы: апиология.


Глобальное сокращение численности насекомых-опылителей стало одной из самых серьезных экологических и сельскохозяйственных проблем нашего времени. Это явление представляет угрозу не только для биоразнообразия, но и для продовольственной безопасности, так как многие сельскохозяйственные культуры зависят от опыления насекомыми (Potts et al., 2016; Powney et al., 2019). Среди всех опылителей медоносная пчела (Apis mellifera) является ключевым видом, используемым как в естественных, так и в управляемых экосистемах. Однако за последние десятилетия численность колоний A. mellifera сокращалась (Goulson et al., 2022; Siviter et al., 2021), что вызывает серьезные опасения в научном сообществе (Vanbergen, 2013; VanEngelsdorp et al., 2009).
Эктопаразитический клещ Varroa destructor был признан как один из главных факторов, способствующих снижению численности пчелиных семей (Rosenkranz et al., 2010). Этот паразит наносит ущерб хозяевам как непосредственно, питаясь их тканями, так и косвенно, передавая вирусы, например вирус деформации крыльев DWV (Nazzi et al., 2012; Wilfert et al., 2016). Другой опасный эктопаразитический клещ, Tropilaelaps mercedesae, также поражает семьи пчел A. mellifera. Этот вид клещей встречается в Азии и может сосуществовать с V. destructor в одном улье (Anderson & Roberts, 2013). Хотя клещи T. mercedesae не так широко распространены, как V. destructor, их способность к более быстрому размножению и сокращенная стадия развития делают их потенциально более опасными паразитами медоносных пчел (Chantawannakul et al., 2018). Кроме того, в отличие от V. destructor, T. mercedesae полностью зависят от расплода пчел и не могут питаться на взрослых особях, что связано с морфологическим строением их ротового аппарата (Woyke, 1984; Anderson & Roberts, 2013) (рисунок 1).

Рисунок 1 Фотографии взрослых самок паразитических клещей, снятые с помощью сканирующего электронного микроскопа. Дорсальные (a) и вентральные (b) виды V. destructor сравниваются с дорсальными (c) и вентральными (d) видами T. mercedesae. Оба эктопаразита, паразитирующие на медоносной пчеле A. mellifera, имеют сходства, несмотря на различия в размере и форме тела. Все масштабные линейки — 0,5 мм. Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
Жизненные циклы паразитических клещей тесно связаны с их хозяевами. У V. destructor чередуются стадии: расселения, когда взрослые клещи прикрепляются к пчелам-кормилицам и расселяются, и репродуктивная стадия, проходящая в запечатанных ячейках расплода. На стадии расселения клещи питаются жировым телом взрослых пчел, используя межсегментные мембраны брюшка (Ramsey et al., 2019). Однако во время репродуктивной стадии они переключаются на гемолимфу куколок, что связано с физиологическими особенностями этой стадии развития хозяев (Haunerland & Shirk, 1995).
Хотя гемолимфа исторически считалась основной пищей V. destructor (Rosenkranz et al., 2010), недавние исследования показали, что на стадии расселения клещи предпочитают жировое тело пчел, а не гемолимфу (Ramsey et al., 2019). Это открытие привело к необходимости пересмотра представлений о диетической специализации V. destructor. Более того, остается неясным, как именно стадия жизненного цикла хозяина влияет на выбор источников пищи и метаболические адаптации паразитов.
Клещи V. destructor и T. mercedesae обладают приспособлениями, которые позволяют им эффективно питаться тканями хозяев. У клеща T. mercedesae наблюдается более короткий жизненный цикл, что позволяет им быстрее увеличивать численность популяции в ульях (Anderson & Roberts, 2013). Эти различия между двумя видами требуют дополнительных исследований, чтобы понять, как стадии жизненного цикла хозяев влияют на их паразитов и какие метаболические адаптации необходимы для использования различных источников пищи.
Гипотеза авторов заключается в том, что V. destructor и T. mercedesae демонстрируют диетическую специализацию, зависящую от стадии жизненного цикла медоносных пчел. Han et al. (2024) предположили, что репродуктивные самки клещей V. destructor питаются преимущественно гемолимфой куколок, а расселяющиеся клещи V. destructor предпочитают жировое тело взрослых пчел, тогда как клещи T. mercedesae, полностью зависят от расплода пчел и демонстрируют уникальную диетическую специализацию.
Для проверки своей гипотезы Han et al. (2024) провели серию экспериментов, включающих витальное окрашивание, протеомный и метаболомный анализы, а также морфологическое изучение клещей. Исследование проводилось на восьми семьях медоносных пчел (Apis mellifera) с естественным заражением клещами V. destructor и T. mercedesae. Семьи содержались на пасеке Института исследований пчеловодства Китайской академии сельскохозяйственных наук. Все семьи поддерживались в стандартных условиях, не подвергались обработке акарицидами, чтобы сохранить естественный уровень зараженности клещами.
Для определения предпочтительных мест питания клеща V. destructor были отобраны куколки пчел с розовыми и пурпурными глазами (оптимальные стадии развития расплода). Куколки извлекались из ячеек и окрашивались трипановым синим для визуализации ран от проколов, нанесенных клещами. Местоположения ран фиксировались с использованием стереомикроскопа Leica EZ4W. Для анализа использовались более 1200 куколок, чтобы определить распределение мест прокусывания клещами.
Для выявления источников пищи клещей проводилось флуоресцентное окрашивание тканей куколок пчел: гидрофильный уранин использовался для окрашивания гемолимфы (Kanost, 2009; липофильный нильский красный применялся для маркировки жирового тела (Chen et al., 2009).
Куколок погружали в растворы красителей, чтобы обеспечить равномерное распределение меток. Окрашенные куколки помещались в инкубатор, где их заражали V. destructor и T. mercedesae. После кормления клещи извлекались для анализа под флуоресцентным микроскопом. Количественная оценка флуоресцентных сигналов уранина и нильского красного проводилась для определения, какая ткань была потреблена.
Для анализа потребляемых тканей клещей использовали масс-спектрометрию. Белки извлекали из клещей, собранных на стадии размножения (из куколок) и на стадии расселения (из взрослых пчел). Идентификация белков проводилась с использованием референсных протеомных баз данных медоносных пчел. Основное внимание уделялось белкам, специфичным для гемолимфы и жирового тела, таким как MRJP и гексамерины (Martins et al., 2008).
Метаболомное профилирование проводилось для изучения метаболических различий между стадиями жизненного цикла клещей. Образцы клещей (протонимфы, дейтонимфы, репродуктивные и расселяющиеся самки) собирали и замораживали при -80°C. Для анализа использовались методы газовой и жидкостной хроматографии, сопряженной с масс-спектрометрией (GC-MS и LC-MS). Определялись метаболиты, связанные с белковым и липидным обменом.
В результате был осуществлен всесторонний анализ пищевых предпочтений клещей Varroa destructor и Tropilaelaps mercedesae в зависимости от стадии жизненного цикла их хозяев. Анализ 1285 куколок пчел показал, что 98,5% клещей V. destructor создавали только одну рану для своего питания, и более 85% этих ран располагались на втором брюшном стерните. Остальные проколы были обнаружены на мезотораксе (10%) или других сегментах брюшка (4,3%) (Donzé & Guerin, 1994). Такое предпочтение второго брюшного стернита объясняется его тонкой кутикулой, что облегчает доступ к гемолимфе. Это место также обеспечивает клещам устойчивую среду для продолжительного питания и минимизацию энергетических затрат (Kanbar & Engels, 2004). Han et al. (2024) предположили, что выбор данного места питания связан с его высокой доступностью и значительным количеством гемолимфы, что подтверждает гипотезу о его адаптивной роли для клеща V. destructor (рисунок 2).
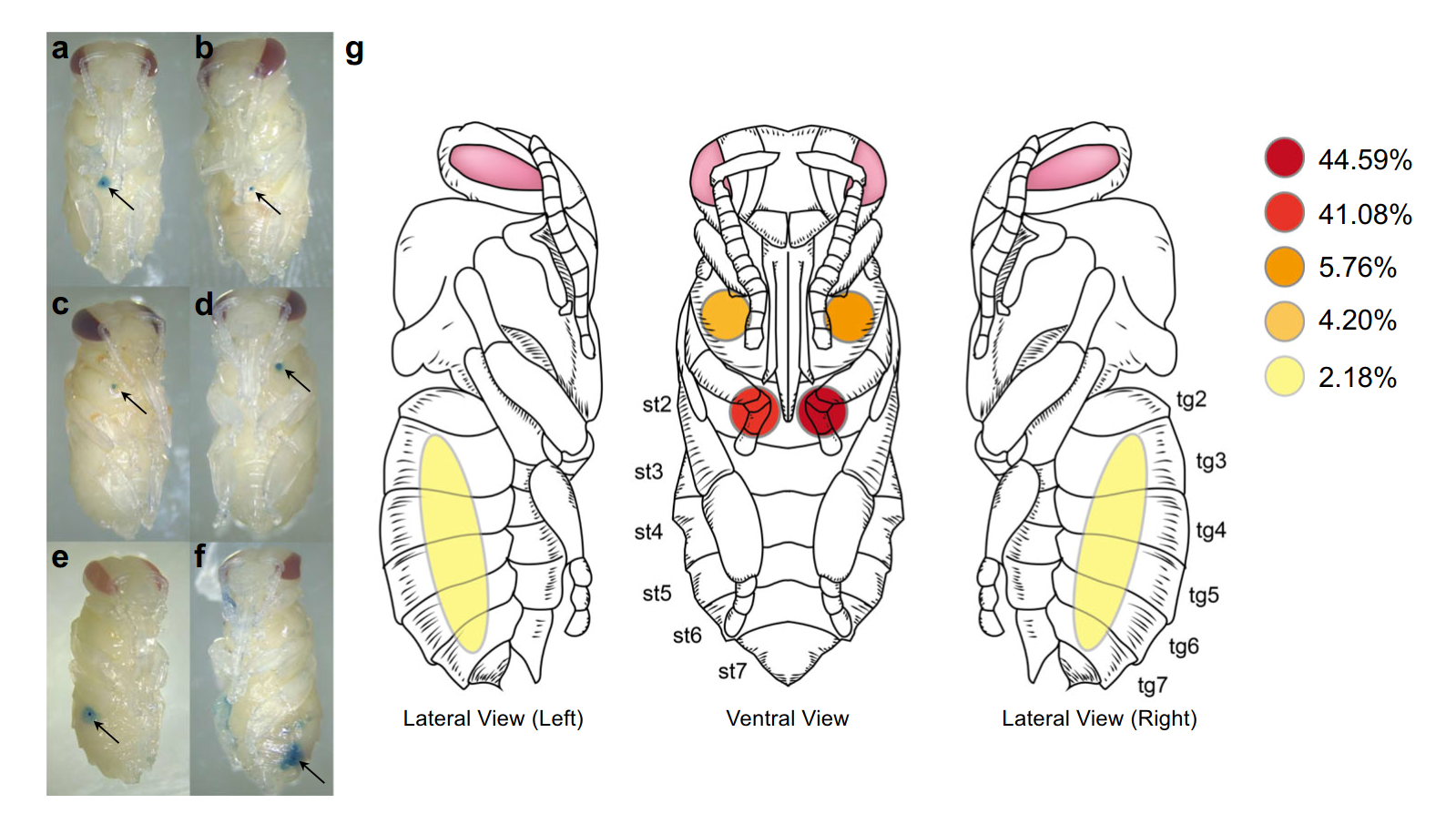
Рисунок 2 Клещ V. destructor демонстрирует предпочтение к питанию на втором брюшном сегменте куколок медоносной пчелы. (a–f) Репрезентативные фотографии различных мест питания V. destructor, выявленных с помощью витального окрашивания повреждений покровов в разных частях куколки. Каждая стрелка указывает на место перфорации клещом. (g) Диаграмма частоты повреждений покровов на 1285 куколках, обследованных восьмью испытаниями (st — стернит, tg — тергит). Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
Клещи T. mercedesae демонстрировали более широкое распределение мест питания на куколках, включая антенны, ноги и хоботок. Тем не менее, как и V. destructor, они преимущественно потребляли гемолимфу, что подтверждено витальным окрашиванием. Han et al. (2024) отметили, что высокая гибкость T. mercedesae в выборе мест питания может быть связана с их коротким периодом стадии расселения и высокой скоростью размножения. Это делает T. mercedesae более зависимыми от гемолимфы куколок пчел, чем V. destructor, который способен паразитировать на взрослых пчелах.
Витальное окрашивание продемонстрировало, что V. destructor и T. mercedesae преимущественно потребляли гемолимфу, что подтверждено преобладанием зеленой флуоресценции (гидрофильного уранина) над красной (липофильного нильского красного) (рисунок 3). У репродуктивных самок V. destructor соотношение уранина/нильского красного составило 1689,81 ± 114,06, значительно превышая показатели у куколок пчел (соотношение 10,19 ± 0,69). В стадии расселения клещ V. destructor демонстрировал противоположные предпочтения, потребляя в основном жировое тело через межсегментные мембраны брюшка. Han et al. (2024) отметили, что различия в рационе клещей связаны с физиологическими особенностями тела хозяина. У взрослых пчел жировое тело сконцентрировано в виде слоя под кутикулой, тогда как у куколок оно распределено диффузно, что делает гемолимфу более доступной (Haunerland & Shirk, 1995). Эти результаты согласуются с выводами Ramsey et al. (2019), где питание гемолимфой наблюдалось как преобладающее на стадии расплода.
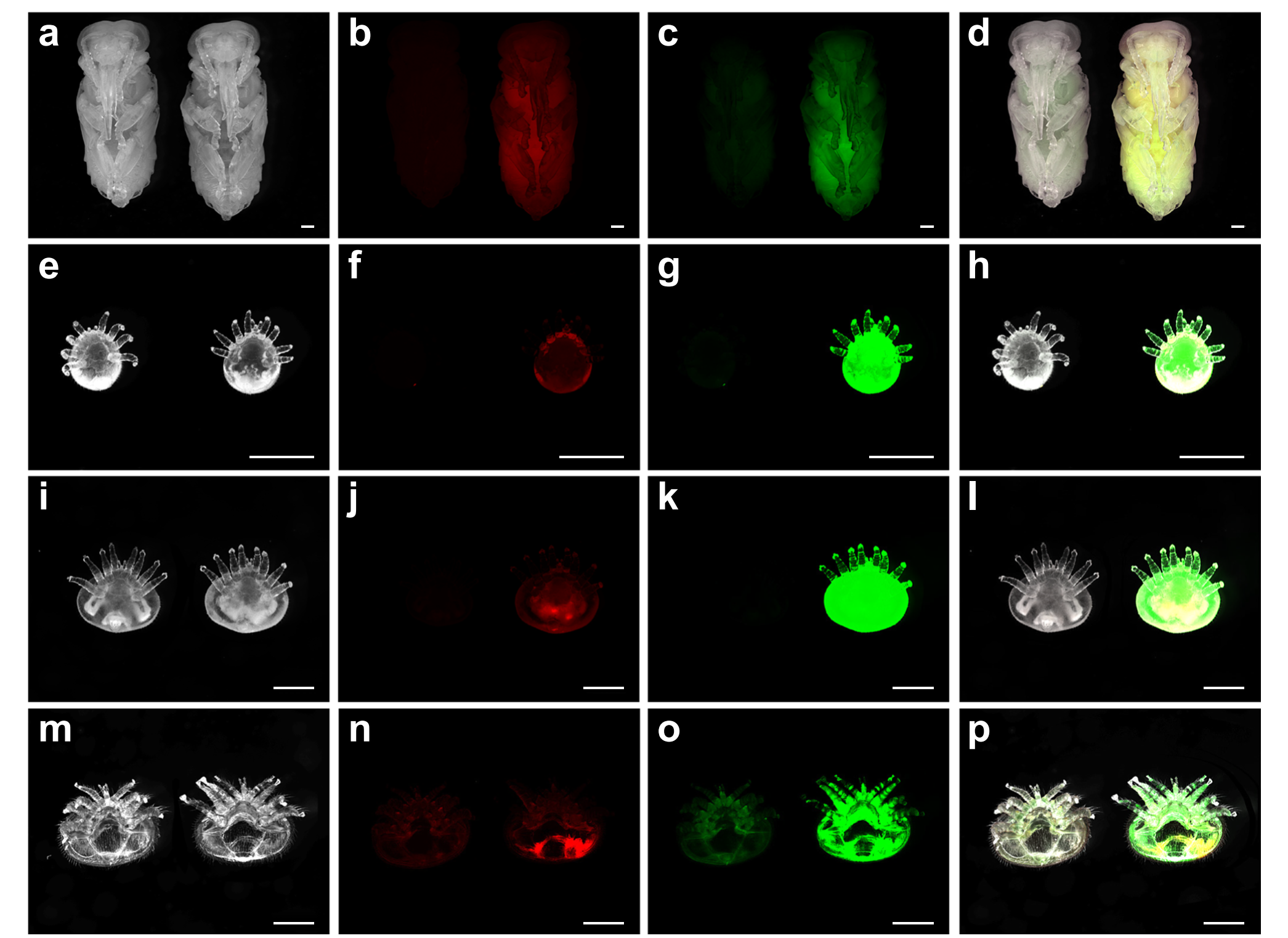
Рисунок 3. Varroa destructor в репродуктивной стадии поглощает большое количество гемолимфы куколок медоносных пчел. Фотографии показывают витально окрашенных куколок, и клещей, питающихся этими окрашенными куколками. Изображения включают светлое поле, флуоресценцию с нильским красным (жировое тело) и уранином (гемолимфа), а также объединенные изображения. На фотографиях слева — контроль, справа — окрашенные образцы. Масштаб 1 мм. Фотографии включают куколок медоносных пчел (a–d), протонимф (e–h), дейтонимф (i–l) и репродуктивных самок (m–p). Каждая строка — не менее 60 особей из шести экспериментов. Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
У репродуктивных самок V. destructor было идентифицировано 272 белка медоносных пчел, из которых 74,2% были белками гемолимфы (Hu et al., 2016). У расселяющихся клещей было обнаружено только 182 белка, из которых 33,3% были белками гемолимфы пчел, что указывает на их переключение на жировое тело. Ключевые белки гемолимфы, ассоциированные с гемолимфой куколок, в частности, MRJP (Major Royal Jelly Proteins), были наиболее представлены у репродуктивных самок. Гексамерины - белки, которые синтезируются жировым телом личинок пчел также присутствуют в гемолимфе. Их концентрация была выше у репродуктивных самок клещей, чем у расселяющихся. Протеомные данные подтвердили переключение клеща V. destructor с гемолимфы на жировое тело в зависимости от стадии жизненного цикла. Более высокое содержание MRJP у репродуктивных самок клещей свидетельствует о том, что гемолимфа куколок пчел является более питательной средой для синтеза белков, необходимых для размножения. Это также подчеркивает их адаптацию к репродуктивной стадии хозяина, где запечатанные ячейки создают защищенную среду для эффективного питания (Donzé & Guerin, 1997) (рисунок 4).
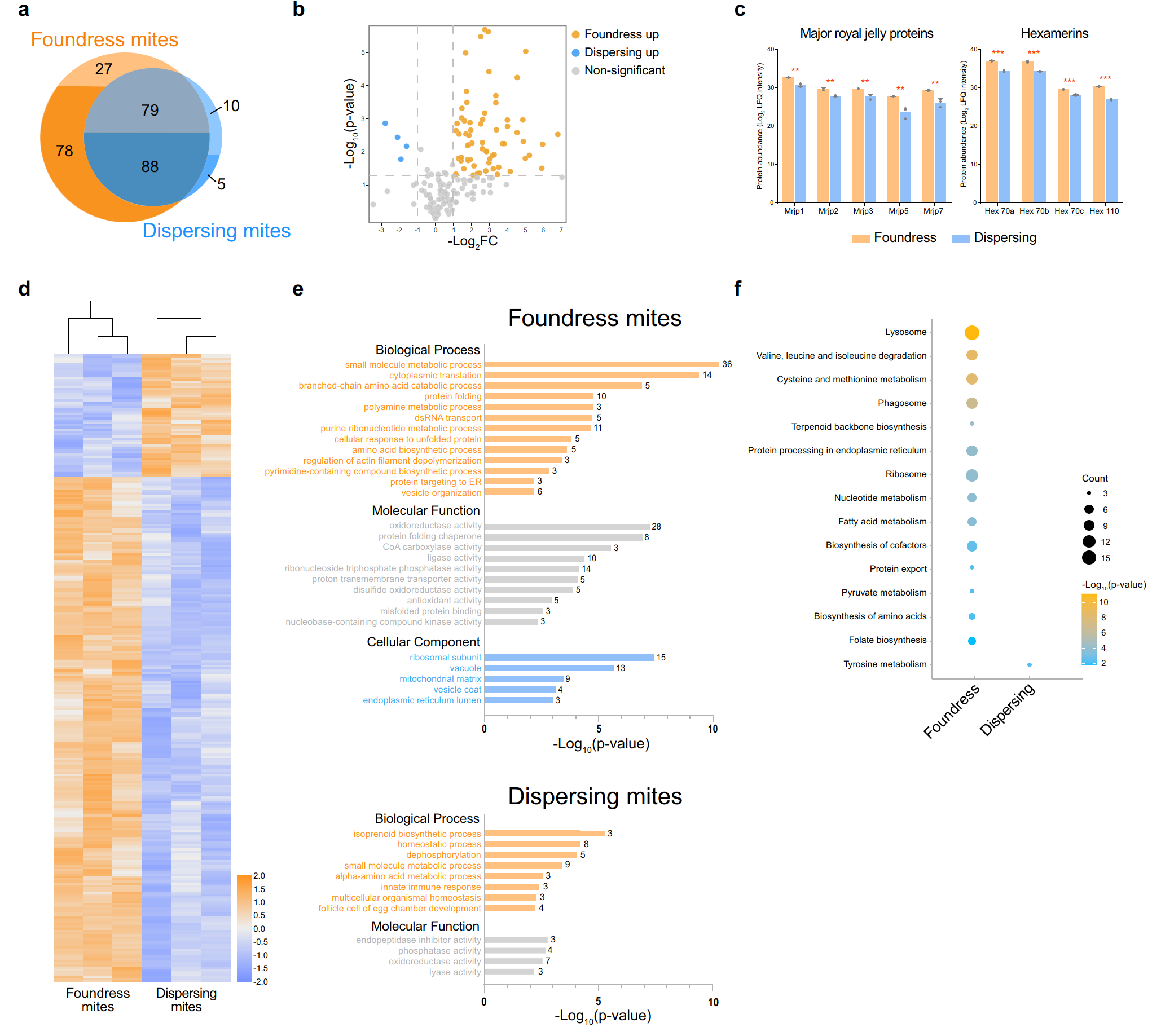
Рисунок 4. Протеомные профили клещей V. destructor у репродуктивных самок и расселяющихся клещей: (a) Большинство белков медоносных пчел, идентифицированных у V. destructor, были найдены в обеих стадиях, но 41,8% были уникальны для репродуктивных самок (питающиеся на куколках) или расселяющихся клещей (питающиеся на взрослых пчелах). Белки, идентифицированные в гемолимфе пчел, чаще встречались у репродуктивных самок. (b) Из 167 перекрывающихся белков 69 различались по количеству: 65 были наиболее представлены у репродуктивных самок, 4 — у расселяющихся клещей. (c) Белки маточного молочка и гексамерины были более многочисленны у репродуктивных самок. (d) Дифференциально экспрессируемые белки были сгруппированы: большая группа у репродуктивных самок клещей, меньшая — у расселяющихся клещей. (e) GO-анализ показал разнообразие биологических функций у разных стадий клещей. (f) Анализ путей KEGG показал более активный белковый метаболизм у репродуктивных самок. Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
Метаболомный анализ выявил существенные различия в метаболических профилях репродуктивных и расселяющихся клещей. У репродуктивных самок было обнаружено обогащение путей, связанных с метаболизмом аминокислот (биосинтез аминоацил-тРНК и метаболизм D-аминокислот), что подтверждает усиленную потребность в белках для размножения. У расселяющихся клещей профиль метаболитов был связан с липидным обменом, включая пути метаболизма глицерофосфолипидов и биосинтеза ненасыщенных жирных кислот. Различия в метаболических профилях согласуются с изменениями в рационе питания. Гемолимфа, потребляемая репродуктивными самками, богата аминокислотами, которые используются для синтеза белков, необходимых для яйцекладки. Липиды жирового тела, потребляемые расселяющимися клещами, обеспечивают высокую энергетическую отдачу, необходимую для длительного прикрепления к взрослым пчелам и избегания их защитных реакций (рисунок 5).
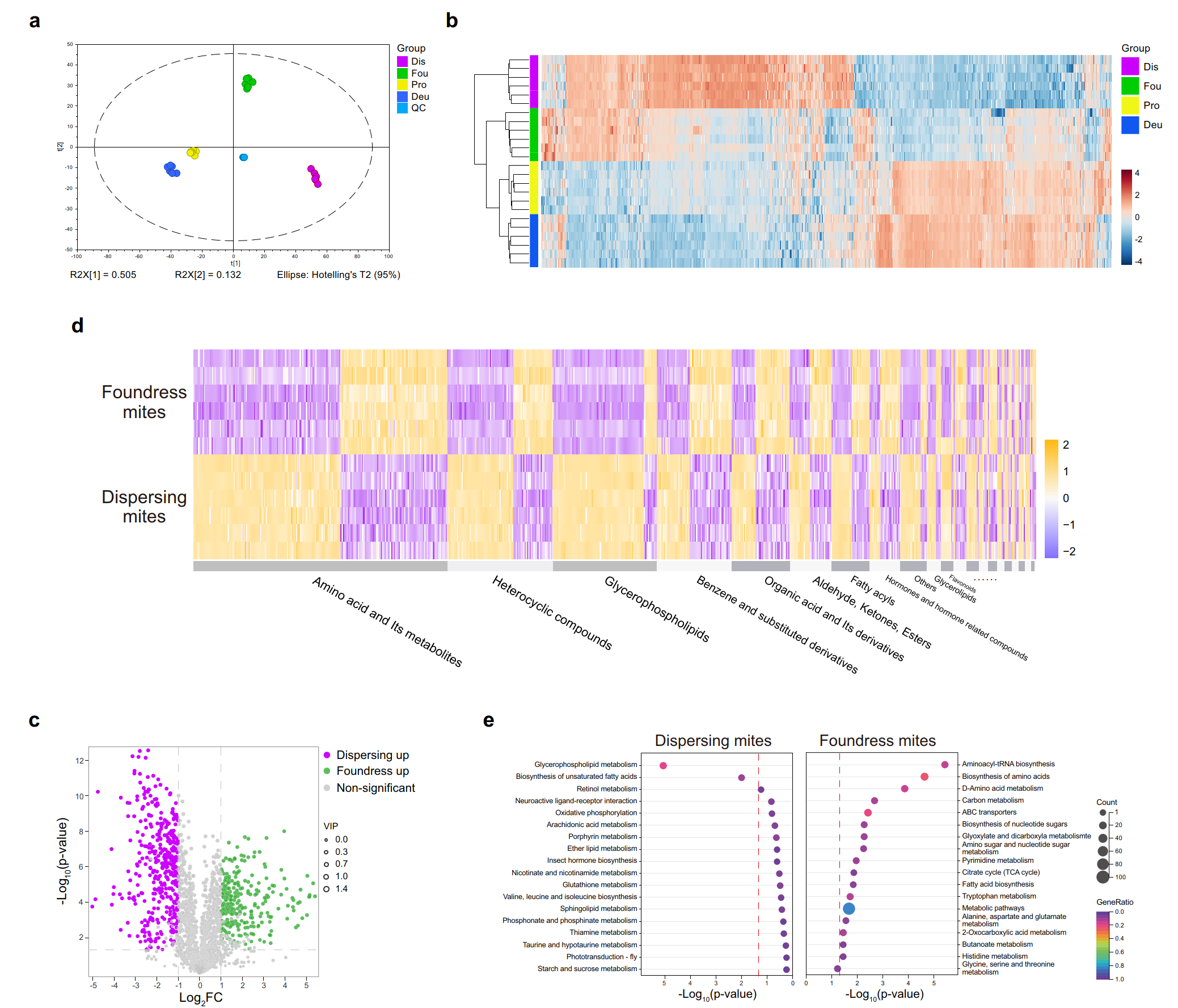
Рисунок 5. Метаболомное профилирование V. destructor (протонимфы, дейтонимфы, репродуктивные самки и расселяющиеся клещи) показывает различия в источниках пищи между репродуктивными самками и расселяющимися клещами. (a) PCA-график метаболитов демонстрирует четкое разделение стадий с шестью повторностями каждой. (b) Тепловая карта и кластеризация метаболитов показывают сходство репродуктивных самок с нимфами по диете, а не возрасту. (c) Вулканный график показывает, что 29,5% метаболитов более обильны у репродуктивных самок (зеленый) и расселяющихся клещей (фиолетовый). (d) Тепловая карта метаболитов между репродуктивными самками и расселяющимися клещами показывает сложную картину. (e) Обогащение путей KEGG показывает более активный белковый метаболизм у репродуктивных самок и более выраженный липидный метаболизм у расселяющихся клещей. Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
Результаты работы Han и соавторов (2024) демонстрируют уникальную диетическую специализацию клещей V. destructor и T. mercedesae. У клеща V. destructor стадия расселения связана с питанием жировым телом взрослых пчел, тогда как стадия размножения – с гемолимфой куколок. Эти паразиты чередуют питание гемолимфой и жировым телом в зависимости от стадии жизненного цикла хозяина. T. mercedesae полностью зависят от гемолимфы и показывают более узкую специализацию. Данные Han et al. (2024) подчеркивают важность адаптивной стратегии паразитов для выживания и успеха размножения. Результаты Han et al. (2024) открывают перспективы для разработки методов борьбы с паразитическими клещами, основанных на нарушении их питания на разных стадиях развития.
Список литературы
- Anderson, D. L., & Roberts, J. M. K. (2015). Standard methods forTropilaelapsmites research. Journal of Apicultural Research, 52(4), 1-16. doi: 10.3896/ibra.1.52.4.21
- Chantawannakul, P., Ramsey, S., vanEngelsdorp, D., Khongphinitbunjong, K., & Phokasem, P. (2018). Tropilaelaps mite: an emerging threat to European honey bee. Curr Opin Insect Sci, 26, 69-75. doi: 10.1016/j.cois.2018.01.012
- Chen, W., Zhang, C., Song, L., Sommerfeld, M., & Hu, Q. (2009). A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. J Microbiol Methods, 77(1), 41-47. doi: 10.1016/j.mimet.2009.01.001
- Donzé, G., & Guerin, P. M. (1994). Behavioral attributes and parental care of Varroa mites parasitizing honeybee brood. Behavioral Ecology and Sociobiology, 34(5), 305-319. doi: 10.1007/bf00197001
- Donzé, G., & Guerin, P. M. (1997). Time-activity budgets and space structuring by the different life stages ofVarroa jacobsoni in capped brood of the honey bee, Apis mellifera. Journal of Insect Behavior, 10(3), 371-393. doi: 10.1007/bf02765605
- Goulson, D., & Nicholls, E. (2022). Anthropogenic influences on bee foraging. Science, 375(6584), 970-972. doi: 10.1126/science.abn0185
- Han, B., Wu, J., Wei, Q., Liu, F., Cui, L., Rueppell, O., & Xu, S. (2024). Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat Commun, 15(1), 725. doi: 10.1038/s41467-024-44915-x
- Haunerland, N. H., & Shirk, P. D. (2009). Regional and functional differentiation in the insect fat body. In V. H. Resh & R. T. Cardé (Eds.), Encyclopedia of Insects (pp. 446-449): Academic Press.
- Hu, H., Bienefeld, K., Wegener, J., Zautke, F., Hao, Y., Feng, M., . . . Li, J. (2016). Proteome Analysis of the Hemolymph, Mushroom Body, and Antenna Provides Novel Insight into Honeybee Resistance against Varroa Infestation. Journal of Proteome Research, 15(8), 2841-2854. doi: 10.1021/acs.jproteome.6b00423
- Kanbar, G., & Engels, W. (2004). Number and position of wounds on honey bee (Apis mellifera) pupae infested with a single Varroa mite. European Journal of Entomology, 101(2), 323-326. doi: 10.14411/eje.2004.041
- Kanost, M. R. (2009). Hemolymph. In V. H. Resh & R. T. Cardé (Eds.), Encyclopedia of Insects (pp. 446-449): Academic Press.
- Martins, J. R., Nunes, F. M. F., Simões, Z. L. P., & Bitondi, M. M. G. (2008). A honeybee storage protein gene, hex 70a, expressed in developing gonads and nutritionally regulated in adult fat body. J. Insect Physiol., 54(5), 867-877. doi: 10.1016/j.jinsphys.2008.03.009
- Nazzi, F., Brown, S. P., Annoscia, D., Del Piccolo, F., Di Prisco, G., Varricchio, P., . . . Pennacchio, F. (2012). Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog, 8(6), e1002735. doi: 10.1371/journal.ppat.1002735
- Potts, S. G., Imperatriz-Fonseca, V., Ngo, H. T., Aizen, M. A., Biesmeijer, J. C., Breeze, T. D., . . . Vanbergen, A. J. (2016). Safeguarding pollinators and their values to human well-being. Nature, 540(7632), 220-229. doi: 10.1038/nature20588
- Powney, G. D., Carvell, C., Edwards, M., Morris, R. K. A., Roy, H. E., Woodcock, B. A., & Isaac, N. J. B. (2019). Widespread losses of pollinating insects in Britain. Nat Commun, 10(1), 1018. doi: 10.1038/s41467-019-08974-9
- Ramsey, S. D., Ochoa, R., Bauchan, G., Gulbronson, C., Mowery, J. D., Cohen, A., . . . vanEngelsdorp, D. (2019). Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc Natl Acad Sci U S A, 116(5), 1792-1801. doi: 10.1073/pnas.1818371116
- Rosenkranz, P., Aumeier, P., & Ziegelmann, B. (2010). Biology and control of Varroa destructor. J Invertebr Pathol, 103 Suppl 1, S96-119. doi: 10.1016/j.jip.2009.07.016
- Siviter, H., Bailes, E. J., Martin, C. D., Oliver, T. R., Koricheva, J., Leadbeater, E., & Brown, M. J. F. (2021). Agrochemicals interact synergistically to increase bee mortality. Nature, 596(7872), 389-392. doi: 10.1038/s41586-021-03787-7
- Vanbergen, A. J., & the Insect Pollinators, I. (2013). Threats to an ecosystem service: pressures on pollinators. Frontiers in Ecology and the Environment, 11(5), 251-259. doi: 10.1890/120126
- Vanengelsdorp, D., Evans, J. D., Saegerman, C., Mullin, C., Haubruge, E., Nguyen, B. K., . . . Pettis, J. S. (2009). Colony collapse disorder: a descriptive study. Plos One, 4(8), e6481. doi: 10.1371/journal.pone.0006481
- Wilfert, L., Long, G., Leggett, H. C., Schmid-Hempel, P., Butlin, R., Martin, S. J., & Boots, M. (2016). Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science, 351(6273), 594-597. doi: 10.1126/science.aac9976
- Woyke, J. (1984). Survival and Prophylactic Control of Tropilaelaps Clareae Infesting Apis Mellifera Colonies in Afghanistan. Apidologie, 15(4), 421-434. doi: 10.1051/apido:198404
Новость подготовили
© д.б.н., в.н.с. Ильясов Рустем Абузарович, Ильясова Алла Юрьевна, Атнагулова Айша Анваровна
11.01.2025