
Говоря о генетических основах видообразования, следует различать две категории признаков – участвующие в формировании адаптаций и предотвращающие свободное скрещивание между особями, обладающими разными адаптациями. Последние могут быть презиготическими и постзиготическими, т.е. проявляющимися у родительских особей при попытке оставить совместное потомство, и реализуемыми уже у самого потомства. Признаки, относящиеся к категории презиготических, представляют особенный интерес. Они формируются под действием полового отбора и направлены на сохранение родительских адаптаций, экономию половых продуктов и максимальную эффективность применения энергии половозрелых особей на воспроизводство очередного поколения. Выбор «свой-чужой» осуществляется именно по этим признакам. Их реализация заключается в распознании брачного партнера, ответе на его ритуальное поведение, успехе и полноценности копуляции или оплодотворении половых продуктов. Вот о том, что это за признаки и что это за отбор такой, по специфическим характеристикам, не связанным с приспособленностью, мы и поговорим.
До последнего времени речь шла о формальном описании таких признаков и их действия в кон- и гетероспецифичных парах, а по результатам анализа межвидовых гибридов – о локализации групп сцепления, определяющих основную межвидовую изменчивость по таким признакам. Тем не менее, все чаще появляются работы, посвященные анализу конкретных мастер-генов, ответственных за значимую долю изменчивости признака и механизмов их реализации на уровне клеток и тканей. Значительные успехи достигнуты в исследованиях, использующих модели развития постериорных лопастей эпандрия и класперов, фланкирующих гоноподы, у самцов дрозофил клады D. simulans. Эпандрий – это девятый тергит (дорсальный склерит предпоследнего сегмента), и выросты на его задней части имеют своеобразную форму шпильки или клюва с разной степенью изогнутости у родственных видов дрозофил (Рис.1).
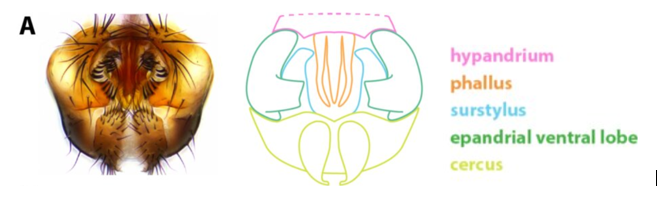
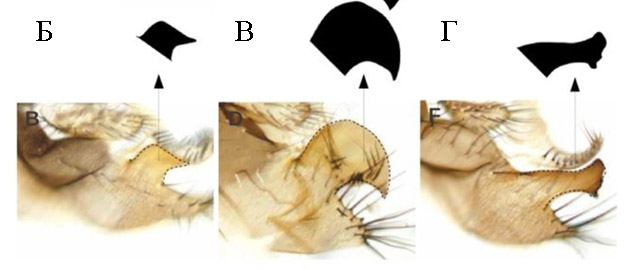
Рисунок 1. Генитальные структуры самцов дрозофил. А - Слева: изображение терминалий взрослого самца D. melanogaster, полученное световой микроскопией. Справа: схема основных терминальных структур. Розовый: гипандрий; оранжевый: фаллос; зеленый: эпандриальная вентральная лопасть; голубой: сурстиль; желтый: церки. Гипандрий выходит за пределы рисунка, как показано пунктирными линиями (из статьи Vincent BJ, Rice GR, Wong GM, Glassford WJ, Downs KI, Shastay JL, Charles-Obi K, Natarajan M, Gogol M, Zeitlinger J, Rebeiz M. An Atlas of Transcription Factors Expressed in Male Pupal Terminalia of Drosophila melanogaster. G3 (Bethesda). 2019 Dec 3;9(12):3961-3972. doi: 10.1534/g3.119.400788. Creative Commons Attribution 4.0 International License); Б, В, Г – микрофотографии эпандриев самцов трех видов комплекса melanogaster: D. melanogaster (В), D. simulans (D) и D. sechellia (F) (из статьи Yassin A, Orgogozo V. Coevolution between Male and Female Genitalia in the Drosophila melanogaster Species Subgroup. PLoS ONE. 2013. 8(2): e57158. https://doi.org/10.1371/journal.pone.0057158, Creative Commons Attribution License).
Эти выросты расположены в месте смыкания эпандрия и гипандрия (вентральной пластины), в непосредственной близости от выдвигающегося фаллоса, и служат для фиксации яйцеклада самки при копуляции. Дистальные боковые части эпандрия, морфологически часто независимые, представлены класперами (или сурстилями) и служат для фиксации всей анальной пластины самки при копуляции.
Но прежде, чем обсуждать генетические основы видоспецифических признаков гениталий, скажем несколько слов об истории формирования научного интереса к этой теме. Различия формы копулятивного аппарата самцов близкородственных видов часто являются основным или даже единственным систематически-значимым признаком. Зоологи-систематики давно подозревали, что ускоренная дивергенция видов по форме гениталий носит неслучайный характер. Дюфур почти 200 лет назад выдвинул гипотезу «ключ-замок», согласно которой соответствие морфологических видовых признаков мужских и женских гениталий является условием успешного спаривания и обеспечивает межвидовую изоляцию. На протяжении ХIХ-XX веков велась дискуссия по поводу справедливости данной гипотезы, и были найдены примеры, подтверждающие действие правила «ключ-замок» у одних видов, и исключающие ее у других. Соответственно, были выдвинуты различные гипотезы об эволюционных механизмах формировании видоспецифических признаков гениталий. Так, Майр считал, что, будучи внутренними структурами, гениталии защищены от действия отбора и их различия вызваны действием плейотропных эффектов генов, участвующих в реализации общих морфологических признаков (Mair, 1963). Гипотезу «ключ-замок» рассматривали с позиции «механического конфликта», связанного с нейтральным накоплением изменчивости по признакам гениталий (Alexander et al., 1996; Arnqvist, 1997).
На эволюционную значимость этих признаков при формировании изолирующих барьеров обратили внимание в 80-е годы ХХ века Койн (Coyne, 1983), Эберхард (Eberhard, 1985, 1988) и Робертсон (Robertson, 1988). Но как действует отбор на морфологические признаки полового аппарата, прямо не связанные с приспособленностью? Когда действует половой отбор на признаки полового аппарата, на прекопуляционной стадии, посткопуляционной или вообще на постзиготической, что заставляет их так быстро меняться? Обсуждая существенные различия в длительности и эффективности спаривания в реципрокных межвидовых спариваниях, авторы предположили ведущую роль морфологических различий копулятивного аппарата самцов родственных видов и действие полового отбора по этим признакам на ранних этапах дивергенции. Последний может быть опосредован отбором по «хорошим генам» или реализуется непосредственно через предполагаемые механизмы «распознания гениталий» (или «выбора самки», или Фишеровского отбора), конкуренции спермы и «конфликта полов», приводящие к коэволюции морфологических структур гениталий самцов и самок. Основываясь на данных, полученных в экспериментах с близкородственными видами дрозофил, Койн и Орр отметили исключительную важность презиготической изоляции и постулировали прямое действие отбора на формирование половой изоляции между дивергирующими видами (Coyne, Orr, 1998).
Роль полового отбора в формировании изоляции между видами на основе несоответствия гениталий самцов и самок долгое время была темой споров (Hosken, Stockley, 2004). В более общем виде спор о механизмах полового отбора продолжался с 1871 г., с книги Чарльза Дарвина «The Descent of Man, and the Selection in Relation to Sex», где Дарвин предположил два основных механизма действия полового отбора – через конкуренцию самцов и через выбор самок. Культурные нормы этого времени неизбежно довлели над частными представлениями большинства ученых, и предположение о выборе самок казалось маловероятным по сравнению с конкуренцией самцов (Huxley, 1938; Hunt et al., 2008 - review). Ситуация меняется к 80-м годам XX века, когда Ланде и Арнольд разрабатывают метод оценки формы и силы полового отбора, основываясь на относительной приспособленности родителей по потомству и на множественной регрессии изучаемых признаков на приспособленность (Lande, Arnold, 1983; Hunt et al., 2007). Используя этот подход и его более совершенные варианты, например, с применением канонического анализа (Phillips, Arnold, 1989; Hunt et al., 2008), для более точной оценки таких форм отбора, как стабилизирующий или дизруптивный, ряд исследований подтверждает действие механизма выбора самок (Andersson, 1994). Более того, удается показать как одновременное, так и последовательное действие механизмов конкуренции между самцами и выбора самок (Hunt et al., 2008).
Возвращаясь к эволюции полового аппарата, следует отметить, что конкуренция между самцами опосредована взаимоотношениями самца и самки, т. е. в итоге – конфликтом полов. Суть этого конфликта заключается в конкуренции над контролем ресурса, вкладываемого родителями в следующее поколение: чей выбор – того и тапки, т. е. тот и обеспечивает наилучшую генетическую среду для своего генотипа. Работы в этом направлении актуальны до сегодняшнего дня. Еще в 2015 г. ЛеВассер-Вьен с соавт. опубликовала статью с говорящим названием «No evidence for external genital morphology affecting cryptic female choice and reproductive isolation in Drosophila» («Нет доказательств того, что морфология внешних половых органов влияет на скрытый выбор самок и репродуктивную изоляцию у дрозофилы»), хотя в этом же исследовании было подтверждено влияние морфологии постериорных лопастей эпандрия на успех спаривания, что прямо указывает на действие прекопулятивного полового отбора. Но исследования продолжались, и доказательства появлялись. Работы Клариссы Хаус с соавт. на D. simulans с использованием линий с различной изменчивостью морфологических характеристик постериорных лопастей эпандрия и методов геометрической морфометрии привели строгое доказательство действия полового отбора на морфологические признаки копулятивного аппарата самцов (House et al., 2013, 2021). Причем было показано действие как стабилизирующего, так и направленного отбора в зависимости от повторности спариваний и конкурентоспособности самцов. В более изощренных генетических экспериментах применяли интрогрессивные линии D. simulans, несущих шесть вариантов участков 2-й хромосомы D. mauritiana (Tanaka et al., 2018). Эти эксперименты подтвердили нарастание механической несовместимости гениталий самца из интрогрессивной линии и самки D. simulans при копуляции в зависимости от отличия признаков копулятивного аппарата от видоспецифичной нормы.
Многочисленные данные, полученные для различных групп насекомых, представлены в обзоре Ли Симмонса (Simmons, 2014), и охватывающие значительно более широкий набор таксонов, от моллюсков до млекопитающих – в обзорах Бреннан с соавторами (Brennan, Prum, 2015; Brennan, Orbah, 2020). Поскольку роль копулятивного аппарата в эволюции видов доказана, мы наконец обратимся к экспериментам, проведенным на близкородственных видах дрозофил с целью выявления генетических основ этих признаков.
Наиболее часто используемой и хорошо изученной является модель сравнения морфологических различий структур генитального комплекса в межвидовых скрещиваниях у близкородственных видов группы melanogaster. Модельные эксперименты применялись для оценки механизмов действия отбора на эффективность производства потомства при межвидовой гибридизации и для поиска генов-мишеней, отвечающих на действие отбора и приводящих к ускоренной эволюции генитальных структур. В экспериментах по скрещиванию D. mauritiana и D. sechellia, значительно отличающихся по форме задних эпандриальных долей у самцов, действие отбора на компоненты приспособленности самцов и самок было связано со значительным травмирующим действием копуляции в гетероспецифических парах, снижением плодовитости и продолжительности жизни самок, уменьшением количества передаваемой спермы (Frazee et al., 2021).
Исследования генетических основ эволюционных изменений лопастей эпандрия дрозофил клады D. simulans были инициированы более четверти века назад в лаборатории Кэтрин Лурье (Liu et al., 1995, 1996; True et al., 1997; Zeng et al., 2000; Tanaka et al., 2018) и показали участие не менее 8 групп сцепления, расположенных на Х-, 2-й и 3-й хромосомах. В последующих исследованиях внимание было уделено одной из наиболее значимых групп сцепления генов, расположенной на левом плече третьей хромосомы, и транскрипционному фактору Poxn, расположенному на второй хромосоме. И если гены, контролирующие развитие генитальных структур и расположенные на 3-й хромосоме, были проанализированы в результате последовательно планируемых исследований, то роль гена Poxn в формировании постериорных лопастей эпандрия была обнаружена случайно.
Транскрипционный фактор Poxn контролирует широкий набор биологических процессов, включая формирование мозга и центральной нервной системы, системы дыхалец, развитие хемосенсорных щетинок и нейронов. В ходе анализа функциональной активности аллелей этого гена, определяющих нарушения формирования хемосенсорных щетинок, была обнаружена стерильность самцов, и анализ ее причин показал нарушение морфологических структур копулятивного аппарата самца, и вызванное этим нарушение брачного поведения (Boll, Noll, 2002). В этой и последующих работах была изучена связь экспрессии Poxn и формы лопастей эпандрия, успеха спаривания самцов и плодовитости оплодотворенных ими самок (Boll, Noll, 2002; Glassford et al., 2015; Frazee, Masly, 2015). Для анализа роли Poxn в контроле формы лопастей эпандрия использовали методы: подавления экспрессии гена с помощью РНК-интерференции (система Gol4-UAS-PoxnIR) ; спасения 0-аллеля Poxn ΔM22-B5 в скрещиваниях с трансгенными мухами с интегрированными конструкциями Poxn-Gol4driver с различными наборами энхансеров из области промотора Poxn; оценки экспрессии репортерного гена из конструкции P{y + UAS-GFP} с упомянутыми драйверными конструкциями Poxn-Gol4driver иммуногистохимического окрашивания белка Poxn в тканях эмбриона и личинки. Авторы подтвердили соответствие признаков формы лопастей эпандрия и характера экспрессии Poxn(Рис. 2), и сопоставляя генотип/фенотип самцов и показатели компонент приспособленности (количество яиц и потомства от данного самца, количество сперматозоидов, переданных от одного спаривания, продолжительность совокупления) показали действие полового отбора на изменчивость данного признака (Рис. 3). Экспрессия Poxn осуществляется под контролем Abd-B в составе Hox- регулируемой сигнальной сети, эволюционно-консервативная функция которой связана с контролем формирования дыхалец в других члениках абдомена. Обобщая полученные результаты, авторы делают вывод о коопции этой сигнальной сети при формировании эволюционно-нового признака постериорных лопастей эпандрия в группе видов melanogaster. В заключении можно только отметить, что во всех работах мишень транскрипционного фактора Poxn осталась неизвестной и путь от этого гена к признаку – не изученным.
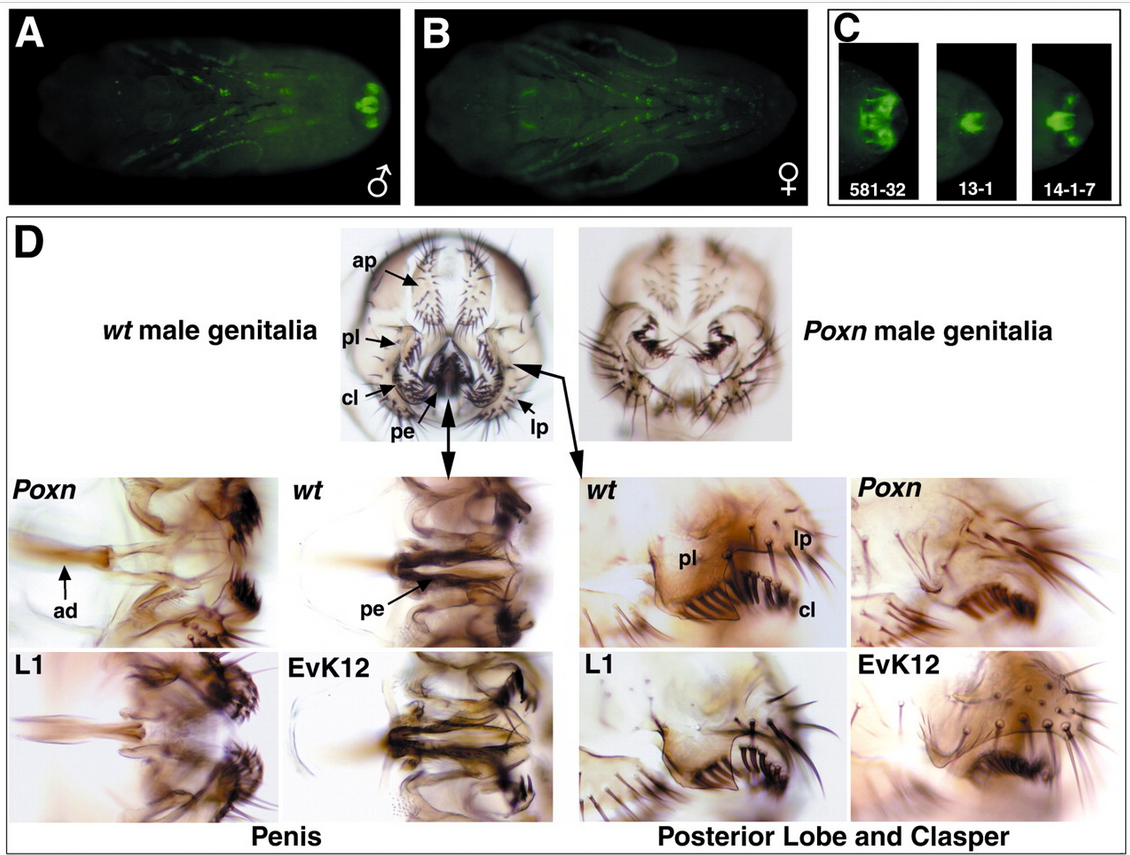
Рисунок 2. Половой диморфизм экспрессии Poxn в куколке в генитальной области и кутикулярный фенотип Poxn ΔM22-B5 мужских гениталий. (A, B) Экспрессия GFP анализируется на вентральной проекции (спереди слева) у самцов (A) и самок (B) w; Куколки Poxn-Gal4-14-1/TM6B (48 часов APF) методом флуоресцентной микроскопии с разрешением 10-кратного увеличения. (C) Детали экспрессии GFP в генитальной области куколок самцов (72 часа APF), управляемой тремя различными конструкциями Poxn-Gal4 (Poxn-Gal4-581, Poxn-Gal4-13-1 и Poxn-Gal4-14-7). (D) Фронтальный вид (дорсальной стороной вверх) препаратов кутикулы гениталий самцов дикого типа (w 1118; слева) и мутанта Poxn (справа) под светлопольной микроскопией с разрешением 8-кратного увеличения. Стрелки указывают анальную пластинку (ap), класпер (cl), латеральную пластинку (lp), пенис (pe) и заднюю долю (pl). На четырех панелях ниже сравниваются детали полового члена (слева), а также области задней доли, класпера и боковой пластинки (справа) у дикого типа (wt) и мутантов Poxn (Poxn ) при увеличении в 2–2,5 раза больше. Стрелки указывают пенис дикого типа (pe) и аподему пениса (ad) у мутанта Poxn без пениса. Четыре панели внизу иллюстрируют при том же увеличении, что и панели выше, спасение у мутантов Poxn полового члена, но не задней доли и класпера, с помощью двух копий EvK, которые включают вышестоящий энхансер гениталий, отсутствующий в L1, и задней доли и класпера, но не пениса, двумя копиями L1, которые включают энхансер интрона гениталий, отсутствующий в EvK. На восьми панелях внизу задняя часть находится справа. (Из статьи Boll W, Noll M. The Drosophila Pox neuro gene: control of male courtship behavior and fertility as revealed by a complete dissection of all enhancers. Development. 2002 Dec;129(24):5667-81. doi: 10.1242/dev.00157. CC BY licence terms with authors retaining copyright)
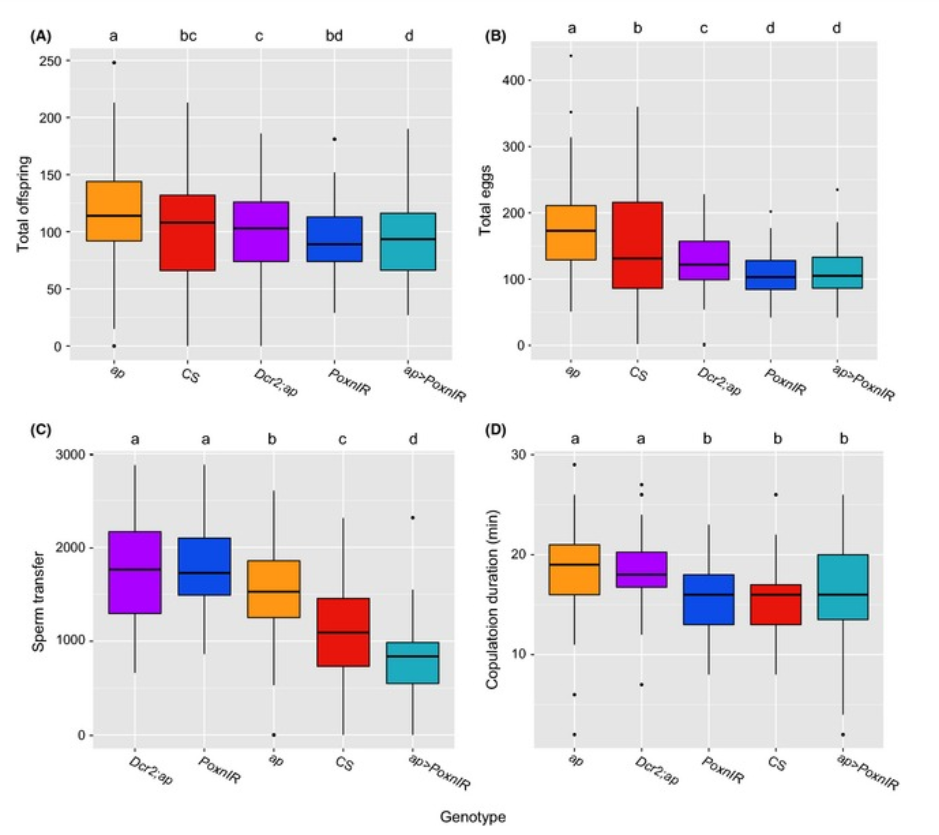
Рисунок 3. Генотипы самцов различаются репродуктивными показателями и реакцией самок на морфологию задней доли. (A) Общее количество потомков, (B) общее количество яиц, (C) количество сперматозоидов, переданных от одного спаривания, (D) продолжительность совокупления. Блок-диаграммы показывают межквартильный размах (IQR), черная горизонтальная линия показывает медиану каждого образца, усы показывают экстремальные значения, а черные точки представляют выбросы. Однородные группы выражены буквами в верхней части рисунков и распределены на основе статистического порога α = 0,05. Сокращения генотипов: ap = ap GAL 4 (контроль с дрожжевым драйвером Gol4); CS = Кантон S (контроль); Dcr2;ap = UAS‐Dcr‐2; ap<>GAL4 (контроль с Dcr‐2 под дрожжевым промотором); PoxnIR = UAS‐PoxnIR (контроль с инвертированными повторами Poxn под дрожжевым промотором); ap>PoxnIR = apGAL4; UAS‐PoxnIR (подавление экспрессии PoxnIR в половом диске). (Из статьи Frazee S.R., Masly J.P. Multiple sexual selection pressures drive the rapid evolution of complex morphology in a male secondary genital structure. Ecology and evolution. 2015, V. 5, N.19, P. 4437-4450. doi:10.1002/ece3.1721; Creative Commons Attribution License).
Очередной цикл статей посвящен локусам, связь которых с терминалиями самцов показана в экспериментах с локализацией генов, контролирующих изменчивость этих структур. В статье Амбер Риджвей с соавт. (Ridgway et al., 2024) показана роль гена Sox21b в формировании изменчивости генитальных структур самцов дрозофил и связанной с этой изменчивостью быстрой диверсификацией видов. Именно эта статья послужила триггером для написания этой новости, или микрообзора. И она, как, впрочем, и все работы, имеет свою предысторию. В 2015 г. группа ученых из Оксфорда под руководством Алистера Макгрегора и Марии Нуньес с привлечением специалистов Института популяционной генетики Вены (Кристиана Шлеттерера) и Университетов Вирджинии и Оклахомы представили результаты QTL-локализации генов, определяющих быструю диверсификацию признаков формы морфологических структур копулятивного аппарата самцов у видов Drosophilasimulans и Drosophila mauritiana . В том числе были идентифицированы локусы на второй хромосоме, правом и левом плечах третьей хромосомы и на Х-хромосоме, связанные с морфологией класперов (структура внешнего полового аппарата самцов, служащая для удержания самок при копуляции), лопастей эпандрия и анальных пластин (рис. 4; Tanaka et al., 2015). Используя известные данные по дифференциальной экспрессии генов между полами (Chatterjee et al. 2011) и между данными видами (Masly et al. 2011), авторы обнаружили 20 генов-кандидатов (Cpr66D, Mcm7, dally, CG6673, Prm, CG32081, iPLA2-VIA, wls, CG14130, CG14835, CG14838, sulf1, sgl, CG16717, Rac1, msl-3, CG32055, CG32082, Mob2, CG11652), расположенных в районах картированных локусов и участвующих в формировании различий генитальных признаков. Из представленного списка нокдаун-эффекты последних 6-и генов сонаправленны с эффектами интрогрессированных областей и с различиями экспрессии самцов и самок. Интересно, что сходные результаты были получены в экспериментах с линиями D.melanogaster , опубликованными коллегами из Канзасского университета практически в это же время (Hackett et al., 2016). Анализируя рекомбинантное потомство от родительских линий с различающимися характеристиками формы задних лопастей эпандрия, авторы отметили четыре локуса на Х-хромосоме, второй хромосоме и обеих плечах третьей хромосомы (Рис. 4), включающие набор генов-кандидатов, изменчивость которых определяет вариации изученных признаков. Более того, состав предполагаемого набора генов хорошо совпадал с представленными в работе Танаки с соавт. (Tanaka et al., 2015).

Рис. 4. Четыре QTL, картированные для значений первой оси принципиальных компонентов (PC1) признаков формы задней лопасти. Две сплошные кривые представляют свидетельства картирования QTL. Фиолетовая кривая получена на основе набора данных с использованием одной случайно выбранной доли от каждого из рекомбинантов, при этом обе доли были успешно оценены, а у остальных мух присутствовала единственная доля. Зеленая кривая использует другую долю для 58% дрозофил, у которых были оценены обе доли. Горизонтальные пунктирные линии представляют собой полученные в результате перестановки статистические пороги 5% по всему геному (X; LOD = 2,73, аутосомы; LOD = 3,57). Положения маркеров представлены в виде делений вдоль внутренней оси X. Интервалы поддержки 2-LOD для каждого QTL выделены серыми полосами. (Из статьи Hackett JL, Wang X, Smith BR, Macdonald SJ (2016) Mapping QTL Contributing to Variation in Posterior Lobe Morphology between Strains of Drosophila melanogaster. PLoS ONE 11(9): e0162573. https://doi.org/10.1371/journal.pone.0162573,; Creative Commons Attribution License).
Дальнейшая работа группы исследователей из Оксфорда была посвящена детальному анализу найденных и поиску новых генов-кандидатов, подтверждению их роли в формировании генитальных структур и определению генетических механизмов изменения этой роли. Так, изучая влияние гена tartan на изменение морфологии класперов (Hagen et al., 2019), были использованы методы картирования с высоким разрешением, анализа транскриптома, нокдауна всех генов-кандидатов с помощью РНК-интерференции, РНК-гибридизации in-situ. На базе интрогрессивных линий с заменой маленькой области на 3-й хромосоме D.simulans в пределах 500 кб в районе гена tartan на фрагмент хромосомы
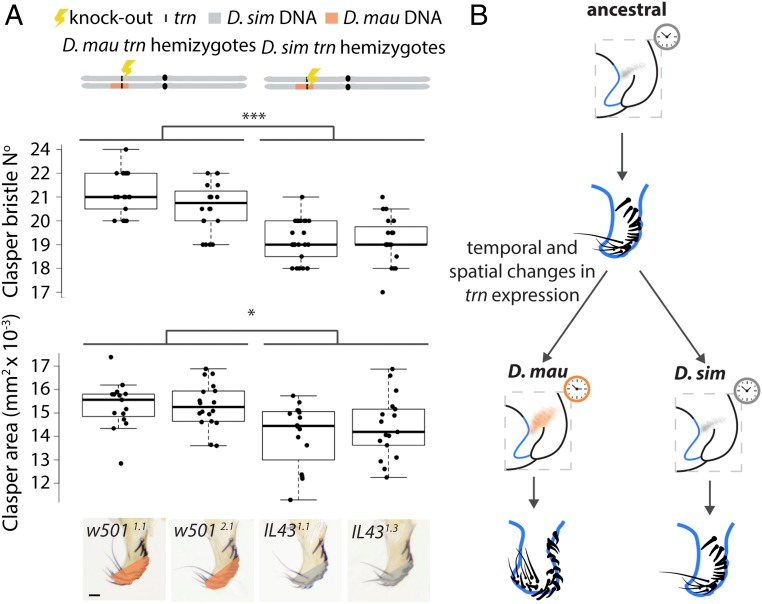
Рисунок 5. Реципрокные гемизиготы trn показывают, что этот локус способствует эволюционным различиям в морфологии мужских класперов. ( A ) Схема вверху иллюстрирует третью хромосому реципрокных гемизигот, несущих функциональный аллель trn только от Dmau D1 ( слева ) и Dsim w 501. Мы обнаружили значительную разницу в площади их застежек ( F (3, 61) = 7,012, P <0,001) и количестве щетинок застежки ( F (3, 83) = 26,29, P <0,001), показанных на диаграммах ниже. Мухи с функциональным аллелем trn D. mauritiana ( IL43 1,1 и IL43 1,3 ) имеют значительно более крупные класперы (* P < 0,05) с большим количеством щетинок (*** P < 0,001), чем мухи с функциональным аллелем trn D. simulans , w501. 1.1 и w501 2.1. В прямоугольниках показаны диапазон, верхний и нижний квартиль, а также медиана для каждого образца. ( B ) Эволюционные изменения увеличили пространственную область и временную экспрессию trn во время развития класперов у D. mauritiana, что привело к появлению более крупных класперов с большим количеством щетинок у этого вида по сравнению с D. simulans . Оранжевая и серая штриховка указывают на широкую и узкую экспрессию trn у основания развивающихся класперов у D. mauritiana и D. simulans соответственно. Часы соответствующего цвета указывают на различия в устойчивости этого домена экспрессии. (Шкала, 20 мкм.) (Из статьи Hagen JFD, Mendes CC, Blogg A, Payne A, Tanaka KM, Gaspar P, Figueras Jimenez J, Kittelmann M, McGregor AP, Nunes MDS. tartan underlies the evolution of Drosophila male genital morphology. Proc Natl Acad Sci U S A. 2019 Sep 17;116(38):19025-19030. doi: 10.1073/pnas.1909829116. Epub 2019 Sep 4. PMID: 31484761; PMCID: PMC6754542. Creative Commons Attribution-NonCommercial-NoDerivatives License 4.0 (CC BY-NC-ND)
Очередная работа авторов, вышедшая в 2021 г., была посвящена поиску новых генов-кандидатов, участвующих в формировании межвидовых различий по показателям класперов и лопастей эпандрия (Hagen et al., 2021). Используя сходный набор методов, более глубокий биоинформатический анализ, в том числе определение ортологов D.melanogaster и анализ генных сетей и генной онтологии, авторы определяют общий набор генов, расположенных в картированных областях, сопоставляют его с дифференциально экспрессирующимися генами у изученных видов и отдельно – с генами, экспрессирующимися в генитальном диске дрозофилы. Последовательно отсекая гены со слабой или не меняющейся (значимо) экспрессией и гены, не экспрессирующиеся в генитальном диске, авторы представляют 12 генов (cuticular protein 66D (Cpr66D), minichromosome maintenance 7 (Mcm7), sugarless [sgl], hairy (h), Grunge (Gug), fear of intimacy (foi), SP1173, CG9953, CG32388, ventral veins lacking [vvl], CG10064, lactate dehydrogenase [ImpL3]), из которых три первых были идентифицированы ранее (Tanaka et al., 2015), но не подтверждены нокдаун-эффектом. В данной работе функциональный анализ этих генов, а также значительного числа экспрессирующихся в генитальном диске в области картирования, был проведен с применением РНК-интерференции на D.melanogaster. В ходе этого анализа подтвердились взаимодействия между генами, в том числе репрессия гена tartan транскрипционным фактором hairy, потенциальная роль гена sugarless в передаче сигналов от Wnt-пути, возможная цепочка регуляций h-Gug-sql. Авторы также делают вывод о возможном механизме смещения области экспрессии целевых генов, при отсутствии различий кодируемых белков и различий в уровнях экспрессии.
Наконец, мы вернулись к статье 2024 г. В последней работе группы из Оксфорда (Ridgway et al., 2024) особое внимание было уделено транскрипционному фактору Sox21b. Развивая ранее полученные результаты, авторы рассматривают дифференциально экспрессирующиеся гены генитального диска D.simulans и D.mauritiana, картированные на левом плече 3-й хромосомы. Используя полученные карты интрогрессии и анализ транскриптома, был продолжен детальный анализ генов, расположенных в районах, связанных с дивергенцией морфологических структур у данных видов. В целевых районах хромосомы располагалось 24 гена, кодирующих транскрипционные факторы, со значительно отличающейся экспрессией между видами. Проверка их функциональной активности с помощью нокдауна на самцах D.melanogaster показала значительное влияние 10 из них на морфологические признаки внешних и внутренних структур копулятивного аппарата. Четыре из них - CKII-a subunit interactor-1, knirps-like, Mediator complex subunit 10 (all cercus area), и Sox21b (posterior lobe area) показали хорошее соответствие эффекта нокдауна и соответствующей фенотипу разнице в экспрессии у D.simulans и D.mauritiana. При этом Sox21b был единственным, расположенным непосредственно в инрогрессированной области. Анализ РНК-гибридизации in-situ на разных стадиях развития герминального диска (рис.6) показал выраженные пространственные различия экспрессии между видами. Sox21b отрицательно регулирует размер задней лопасти эпандрия, и у D.mauritiana отмечена его более выраженная и широкая экспрессия. Проведенный анализ показал отсутствие кооптации Sox21b с Нох-сигнальной сетью, по аналогии с Poxn (Boll, Noll, 2002), потенциально возможное в ходе эволюционных преобразований дыхалец в элементы копулятивного аппарата. Четыре ненейтральные мутации, приводящие к смене аминокислот в составе белка и различающие виды D.simulans и D.mauritiana, не затрагивают активные центры и структурные связи белка, поэтому авторы предполагают наиболее вероятным регуляторную эволюцию белка. Для окончательного подтверждения функциональной роли Sox21b были изготовлены линии с 0-аллелями, используя CRISPR-Cas9 систему и встраивание флуорофора 3XP3-DsRed в экзон 1, что нарушало рамку считывания. Увеличение задней лопасти соответствовало ожидаемому. Затем были подготовлены реципрокные гемизиготы по этому гену, с экспрессией гена одного из родительских видов на фоне генома второго вида. Полученные результаты также соответствовали ожиданию и показали изменение формы и размера лопасти в соответствии с видовой специфичностью экспрессируемого гена. При этом ссаживание пар D.simulans- D.simulans с аллелем Sox21b у самца от D.mauritiana резко сокращал продолжительность копуляции, что подтверждало действие полового отбора на изменчивость признака, связанную с данным геном.
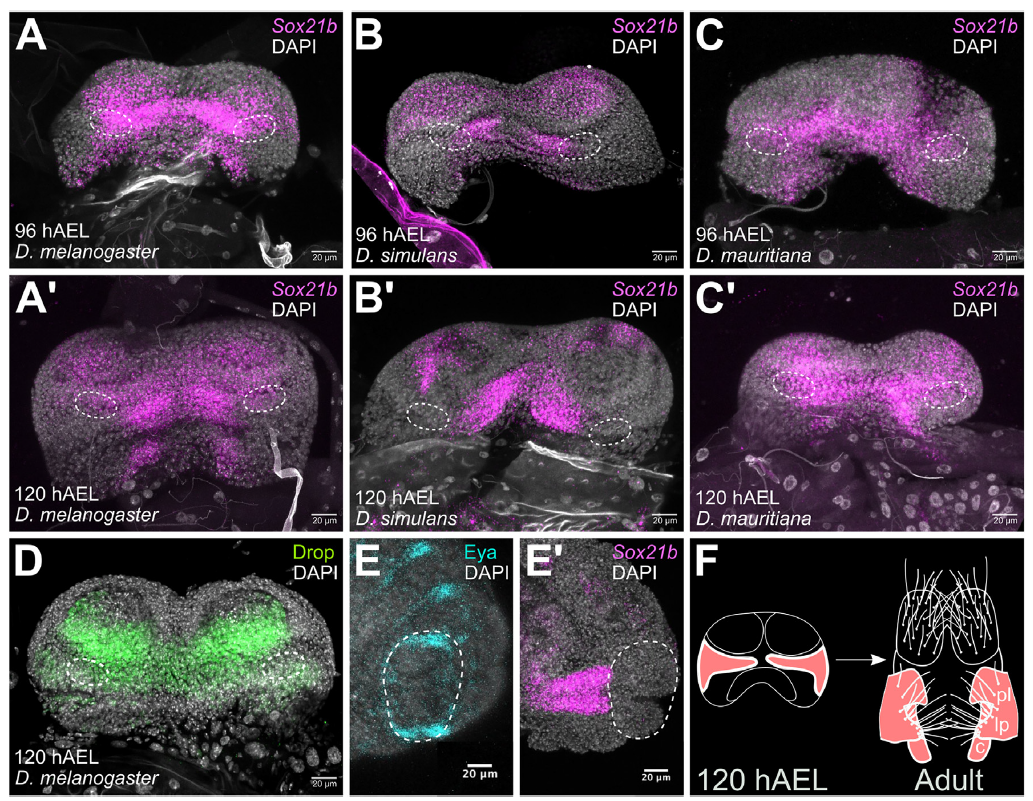
Рисунок 6.Экспрессия Sox21b в генитальных дисках D. melanogaster, D. simulans и D. mauritiana. (A-C') (A-C) Sox21b - цепная реакция гибридизации in situ (HCR) мужских половых дисков при 96 hAEL и (A0-C0) 120 hAEL. (А и А0) D. melanogaster w1118. (B и B0) D. simulans w501. (C и C0) D. mauritiana D1. (A) n = 5, (A0) n = 7, (B) n = 4, (B0) n = 4, (C) n = 4 и (C0) n = 4. Овалы с пунктирными линиями обозначают положение зачатков задней доли. (D) Окрашивание Anti-Drop на мужском генитальном диске D.melanogaster w1118 (n = 8). Положение зачатков задней доли показано овалами с пунктирными линиями. (E и E0) (E) Отсутствие окрашивания Anti-Eyes (Eya) и (E0) Sox21b HCR у эмбриона D. melanogasterw1118 на стадии 16. Развивающиеся задние дыхальца окружены овалами с пунктирными линиями. (Е) n = 15 и (E0) n = 8. (F) Схема, показывающая латеральную пластинку (lp), заднюю долю (pl) и класпер/сурстилюс (c) зачатки у D. melanogaster (Ridgway AM, Hood EJ, Jimenez JF, Nunes MDS, McGregor AP. Sox21b underlies the rapid diversification of a novel male genital structure between Drosophila species. Curr Biol. 2024 Mar 11;34(5):1114-1121.e7. doi: 10.1016/j.cub.2024.01.022. Epub 2024 Feb 2. PMID: 38309269. Creative Commons Attribution license (CC BY 4.0).
Можно отметить, что данный транскрипционный фактор совершенно точно принимает участие в дивергенции видов, обеспечивая в той или иной степени прекопуляционные изолирующие механизмы. Но, еще раз, это транскрипционный фактор .. и пути от гена к признаку лежат через его мишени, так что этот замечательный результат – только дверь на поле возможностей анализа генов-мишеней и разнообразия путей, ведущих от генетической изменчивости к эволюционирующим признакам.
Новость подготовил
© А.М.Куликов
10.07.2024