Часть 3. Стволовые клетки и симбионты.
 © Ересковский Александр Вадимович, ведущий научный сотрудник лаборатории эволюции морфогенезов ИБР РАН. Доктор биологических наук.
© Ересковский Александр Вадимович, ведущий научный сотрудник лаборатории эволюции морфогенезов ИБР РАН. Доктор биологических наук.
Статья международного коллектива биологов развития (Ereskovsky et al., 2022) - третья часть цикла о соматических стволовых клетках у беспозвоночных животных. Этот цикл (Rinkevich et al., 2021; Martinez et al., 2022; Ereskovsky et al., 2022) не только способствует изменению наших представлений о стволовых клетках животных. Он показывает, какую важную роль в изучении этих клеток играют губки (Porifera), и сколько ещё вопросов, касающихся биологии развития Porifera, остаются пока без ответа. Мы надеемся привлечь студентов, аспирантов и исследователей к изучению развития этой группы!
Напомним, что в соответствии с господствующей парадигмой, соматические стволовые клетки (ССК) - коммитированные клоноспецифичные клетки. Их судьбы ограничены тканями/органами, в которых они находятся. Именно они играют ведущую роль в регенерации и восстановлении этих тканей и органов (Clevers, Watt, 2018). Обычно ССК - недифференцированные клетки, которые дают начало либо дочерним стволовым клеткам, либо клеткам, неспособным к самообновлению, либо к клонально-специфическим дифференцированным клеткам. ССК у позвоночных и насекомых обладают высоким ядерно-цитоплазматическим отношением, малы (по сравнению с клонально дифференцированными потомками) и обычно редко встречаются. Однако было показано, что ССК многих водных беспозвоночных очень многочисленны, морфологически разнообразны, демонстрируют широкий спектр состояний дифференцировки. Более того, ССК беспозвоночных могут возникать de novo путем трансдифференцировки из соматических клеток (Rinkevich et al., 2021).
Одним из необычных и пока малоизученных явлений является присутствие внутриклеточных симбионтов в ССК некоторых беспозвоночных. В обзоре (Ereskovsky et al., 2022) представлены данные по ССК и их внутриклеточным симбионтам у губок (Porifera), а также приведено несколько примеров из других групп животных.
Согласно последним исследованиям, у губок существует как минимум четыре типа плюрипотентных ССК: археоциты и хоаноциты (Рис. 1А, В), а также пинакоциты и особые амебоидные вакуолярные клетки (Ereskovsky et al., 2015; Fierro-Constaín et al. 2017; Funayama 2018; Lavrov et al., 2018). Однако сейчас мы уделим внимание археоцитам, поскольку внутриклеточные эукариотические симбионты встречаются только в этих ССК и только у одного класса губок - Demospongiae.
Археоциты представляют собой амебоидные клетки мезохила, лишенные какой-либо полярности или специализированных признаков (Рис 1).Они проявляют высокий полиморфизм и полифункциональность. Что касается функций археоцитов, то они участвуют в транспортировке пищевых частиц и выведении продуктов пищеварения, секреции кремниевых спикул, гаметогенезе и бесполом размножении, иммунитете и т.д. . Такая полифункциональность стволовых клеток необычна для ССК животных.
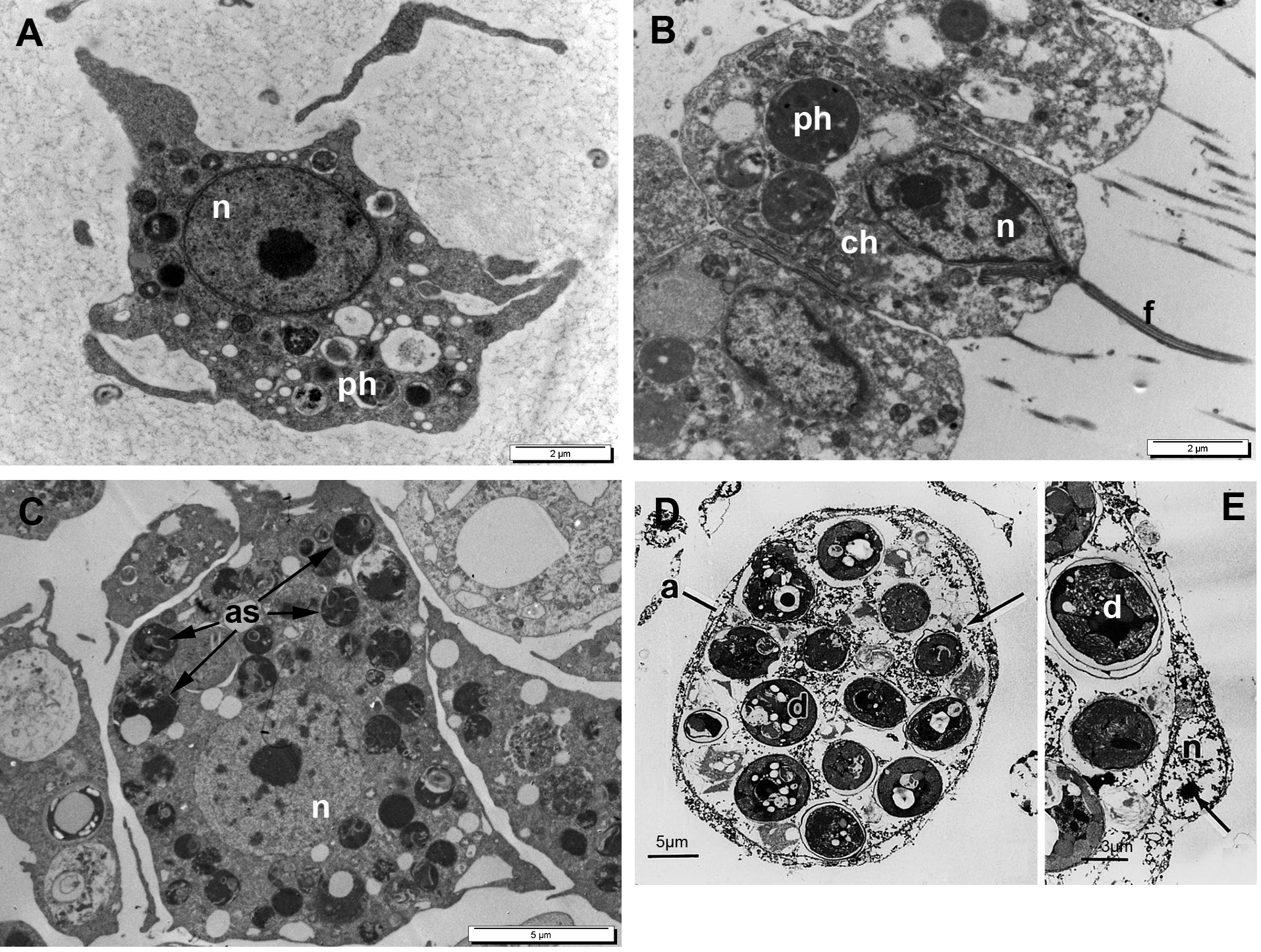
Рис. 1. ТЭМ-фотографии основных плюрипотентных клеток губок. (А) – археоцит Halisarca dujardinii (Demospongiae); (B) — хоаноциты Leucosolenia variabilis (Calcarea); (C) – археоцит пресноводной губки Lubomirskia baicalensis (Demospongiae) с внутриклеточными фотосинтетическими симбионтами; (D, E) – археоциты морской губки Haliclona sp. (Demospongiae) с внутриклеточными симбионтами – динофлагеллятами. as — симбионты - водоросли, ch — хоаноцит, d — динофлагеллята, f — жгутик, n — ядро, ph – фагосома.
Примечательно, что у археоцитов Demospongiae есть еще одна необычная особенность - наличие внутриклеточных фотосинтезирующих симбионтов (Рис. 1С - Е, археоциты с симбионтами). Археоциты пресноводных губок содержат одноклеточные водоросли Chlorophyta из классов Trebouxiophyceae и Chlorophyceae, а также Ochrophyta из класса Eustigmatophytacea. Археоциты некоторых морских демоспонгий включают в себя динофлагеллят Symbiodinium spp. (Zooxanthella). Археоциты обычно включают от трех до 12 симбионтов. В то же время, эндосимбиоз археоцитов и фотосинтезирующих водорослей является факультативным: представители одного и того же вида, обитающие в разных световых условиях могут содержать симбионтов или не иметь их совсем, как, например, в затемненных местообитаниях.
Были проведены эксперименты по заселению симбионтами молодых апосимбиотических губок Ephydatia muelleri. Уже через 4 часа после вылупления из геммул археоциты включали этих симбионтических водорослей (Hall et al. 2021). При половом размножении передача симбионтов происходит горизонтально, то есть из внешней среды. Однако при бесполом размножении за счёт фрагментации или геммулогенеза, передача симбионтов может сочетать как вертикальный перенос, так и горизонтальный.
Роль эндосимбионтов в физиологии губок активно исследуется, но к сожалению, совершенно неизвестно оказывают ли эти фотосинтезирующие симбионты какое-либо влияние на проявление или ингибирование «стволовости» археоцитов.
Интересно, что прокариотические эндосимбионты никогда не обитают в археоцитах или других плюрипотентных клетках губок. Для этого имеются специальные дифференцированные клетки – бактериоциты.
В отличие от демоспонгий, в ССК других многоклеточных животных эндосимбионты встречается редко. Тем не менее, имеется несколько хорошо документированных примеров, которые свидетельствуют о важности ССК в координации и поддержании внутриклеточного симбиоза. Примеры включают, например, глубоководных вестиментифер (Polychaeta), у которых симбиотические бактерии обитают в бактериоцитах. Эти клетки, расположенные в специальном органе — трофосоме, считаются тканеспецифическими монопотентными стволовыми клетками (Pflugfelder et al. 2009). У Hydra (Cnidaria) клоны эпителиальных стволовых клеток (но не интерстициальные клетки), формируют микробные внутриклеточные сообщества (Fraune et al. 2009). У ветвящихся кораллов Stylophora pistillata клетки эндодермы, содержащие водоросли, демонстрируют признаки стволовости. Они экспрессируют гены «стволовости», такие как nanos и tudor, а также гены, продукты которых важны для регуляции клеточного цикла (Levy et al. 2021).
Имеются примеры участия ССК насекомых в поддержании или контроле популяции внутриклеточных симбионтов. На ранних стадиях развития тлей Acyrthosiphon pisum и Megoura viciae, а также таракана Periplaneta americana бактериоциты появляются de novo из апосимбиотических ССК. У P. americana наблюдали также постэмбриональную пролиферацию бактериоцитов. Внутриклеточные симбионты Wolbachia - не единственные цитосимбиотические бактерии в стволовых клетках насекомых. Клетки половой линии также могут быть колонизированы другими микроорганизмами, такими как грамположительная бактерия Spiroplasma у дрозофилы (Hackett et al., 1986) или грамотрицательная бактерия Arsenophonus, которая заражает симбионта Sulcia цикадки Macrosteles laevis (Kobiałka et al. 2016).
До сих пор не известны механизмы, обеспечивающие специфическое поведение ССК в специфических симбиозах. Имеется несколько примеров такого поведения ССК. (1) Непрерывная пролиферация унипотентных бактериоцитов у некоторых кольчатых червей, связанная с образованием новых бактериоцитов. (2) Поддержание симбиоза во время непрерывного образования бактериоцитов из апосимбиотических необластов у плоских червей. (3) Формирование микробных внутриклеточных сообществ эпителиальными стволовыми клетками Hydra. Исследование особенностей функционирования стволовых клеток в этих симбиозах представляет интерес для более глубокого понимания природы ССК Metazoa.
Представленный в этой заметке обзор (Ereskovsky et al., 2022) позволяет взглянуть на ССК беспозвоночных, как на клетки, способные к установлению и поддержанию внутриклеточных симбиозов. Это расширяет наше представление о разнообразии симбиотических ассоциаций, сформировавшихся в ходе диверсификации многоклеточных животных.
Новость подготовил
© Ересковский А.В.
09.05.2022