 © Станислав Кремнёв, кандидат биологических наук, старший научный сотрудник лаборатории эволюции морфогенезов ИБР РАН.
© Станислав Кремнёв, кандидат биологических наук, старший научный сотрудник лаборатории эволюции морфогенезов ИБР РАН.
Научные интересы: морфогенез, эволюция эмбрионального развития.
Нервная трубка в процессе развития позвоночных размечается вдоль дорсовентральной оси на прогенеторные области, из которых в последствии дифференцируются нейроны разных типов. Уникальный набор экспрессии генов в каждом из прогенеторных доменов является результатом тонкой регулировки сигнальных каскадов путем градиентного распределения секретируемых морфогенов. Морфоген Sonic hedgehog (Shh) формирует вентро-дорсальный градиент своего распределения и экспрессируется в клетках дна нервной трубки и хорде. Градиент Shh индуцирует градиентную активность транскрипционных факторов GliA и GliB, которые вместе с Sox2 участвуют в разметке доменов экспрессии специфических генов, отвечающих за дифференцировку разных типов нейронов. Морфогены Wnt и Bmp создают градиент их активности от дорзального полюса нервной трубки и покровной эктодермы.
Дорсовентральная разметка нервной трубки представляет собой классическую модель эмбриональной индукции между тканями. Индукционная активность организатора в данной модели была приписана хорде и сигнальному каскаду shh. Предполагается, что Shh, экспрессия которого начинается в хорде, создает градиент морфогена, что в свою очередь запускает экспрессию Shh в дне нервной трубки, что в свою очередь совместно с другими морфогенами приводит к дорсовентральной разметке нервной трубки.
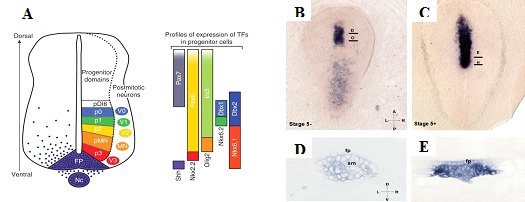
Рис.1 A. Классическая схема индукции shh в клетках дна нервной трубки и ее дорсовентральной разметки (Ribes and Briscoe, 2009). B-E. Пространственная динамика экспрессии shh во время гаструляции кролика. A. Стадия 5-, В. Стадия 5+. D-E. Поперечные срезы. Динамика экспрессии соответствует представлениям классической модели. fp – дно нервной трубки, am – осевая мезодерма
Однако универсальность функции хорды как организатора остается под вопросом. У рыбки Данио ни механическое удаление хорды, ни нарушение формирования хорды у мутантов no tail не приводят к нарушениям дифференцировки дна нервной трубки. У шпорцевой лягушки, активность Shh слабо регулирует экспрессию генов в дне нервной трубки, при этом сильное влияние оказывается на экспрессию латеральных генов, таких как nkx2.2. Также имеются противоречивые данные об индукторе дифференцировки дна нервной трубки у птиц.
Примечательно, что, несмотря на противоречивые данные об источнике сигнала, отвечающего за индукцию дифференцирвки дна нервной трубки, до сих пор не были подробно исследованы начальные стадии экспрессии Shh. Работа Kremnyov et al., 2018, опубликованная в журнале EvoDevo , была направлена на заполнение данного пробела. Исследования показали, что у эмбрионов цыпленка на стадии 5 HH, которые находятся в начале процесса формирования хорды, экспрессия shh выявляется полосой вдоль серединной линии зародыша, впереди гензеновского узелка, а экспрессия в самом гензеновском узелке проявляет лево-правый асимметричный паттерн (рис. 2, А). При подробном исследовании паттерна экспрессии на срезах было показано, что постериорные shh-положительные ткани лежат непосредственно над shh-негативной хордой (рис. 2, D). В серединной области нервной трубки shh-позитивны и осевая мезодерма, и нейроэктодерма, в то время как в антериорной области shh экспрессируется только в прехордальной пластинке, антериорная нейроэктодерма остается негативной. Таким образом, формирующаяся хорда не экспрессирует shh, а экспрессия запускается позже в процессе ее продвижения. Shh в нервной пластинке детектируется на высоком уровне непосредственно в постериорной области, там, где не наблюдается экспресии shh в подлежащей хорде. Такой паттерн экспрессии Shh у курицы ставит под сомнение представления классической модели дорсовентральной разметки нервной трубки.
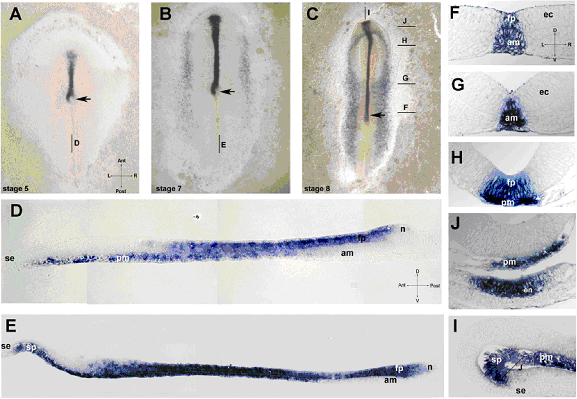
Рис. 2. Пространственная динамика экспрессии shh в развитии курицы. A-C: тотальные препараты эмбрионов на стадии 5 (А), 7 (B) и 8(С); сагиттальный срез эмбриона на стадии 5 (D) и стадии 7 (E); F-J поперечные срезы эмбрионов на стадии 8 HH на уровнях указанных на C. I – сагиттальный срез эмбриона на стадии 8. Обозначения: fp – дно нервной трубки, am – осевая мезодерма, ec – эктодерма, en – эндодерма, n – узелок, pm – прехордальная мезодерма, sp – преоральная область кишки, se – поверхностная эктодерма, стрелка – положение гензеновского узелка, Пересекающиеся стрелки указывают анатомические оси: A – anterior, P – posterior, L – лево, R – право, D – дорзальный, V – вентральный.
Так как shh экспрессируется в презумптивной области дна нервной трубки с самого начала формирования хорды, сразу встаёт вопрос, начинается ли экспрессия shh до формирования гензеновского узелка и прехордальной пластинки. Удивительно, но сигнал shh детектируется уже со стадии 2-3 HH (рис. 3, A). На данной стадии первичная полоска представляет собой сгущение клеточной массы вытянутой в направлении от постериорного полюса к середине бластодиска. Ранее было показано, что область перед первичной полоской на этой стадии является областью презумптивного дна нервной трубки (Lopez-Sanchez et al., 2001). Именно в этой области выявляется сильная экспрессия shh. Также, подробное исследование экспрессии shh на этих стадиях было проведено на срезах (рис. 3, В, С). Полученные данные говорят о том, что экспрессия shh в области дна нервной трубки начинается еще до формирования хорды. Таким образом, классическая модель дорсовентрвльной разметки нервной трубки не может быть применена к развитию птиц в той форме, в которой она представляется в учебниках для всех позвоночных.
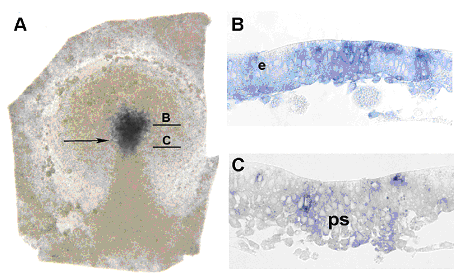
Рис. 3. Ранняя экспрессия shh в эмбрионе курицы на стадиях 2+/3- HH. Стрелкой указана антериорная граница первичной полоски.
Авторы работы также задались вопросом, есть ли вероятность, что функцию разметки нервной трубки у птиц на себя взял другой представитель семейства hedgehog. Оказалось, что нет, экспрессия ни ihh, ни dhh, не была детектирована ни в хорде, ни в дне нервной трубки на исследованных стадиях развития 4- - 17 HH.
Авторами также были проведены сравнительные исследования по выявлению пространственной динамики экспрессии shh у кролика и шпорцевой лягушки. Паттерн экспрессии shh у кролика полностью согласуется с классическими взглядами на дорсовентральную разметку нервной трубки позвоночных (рис 1, B-E), в то время как результаты, полученные на шпорцевой лягушке позволяют предположить индукцию дна нервной трубки в пределах эктодермы в планарной плоскости.
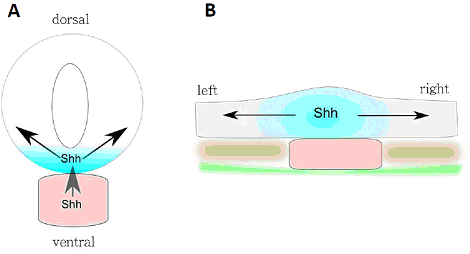
Рис. 4. Схема поперечного среза эмбриона курицы на уровне постериорной хорды. A: вентродорзальная индукция дифференцировки дна нервной трубки согласно принятой модели – shh изначально начинает экспрессироваться в хорде, а далее индуцирует экспрессию shh в дне нервной трубки. Белок Shh из обоих центров формирует градиет. B: изменённая модель демонстрирующая раннюю экспрессию shh в дне нервной трубки и формирование градиента shh в пределах нервной пластинки. Обозначения: розовый – хорда, голубой – дно нервной трубки, зелёный – гипобласт/эедодерма, коричневый - параксиальная мезодерма, стрелки – формирование градиента и индукция.
Сравнительный анализ пространственно-временной последовательности экспрессии shh на окологаструляционных стадиях развития, проведенный в данной работе, имеет большое значение в понимании эволюции разметки нервной системы и будет служить основой для дальнейших исследований функций хорды как эмбрионального организатора. Немаловажно отметить, что модель дорсовентральной разметки нервной трубки где, первопричиной выступает индукция shh от хорды в самых современных статьях и учебных пособиях принимается как данное, однако, результаты, полученные в этой работе, позволяют усомниться в этом и углубить понимание этого важного процесса.
Литература
Kremnyov S., Henningfeld K., Viebahn C., Tsikolia N. Divergent axial morphogenesis and early shh expression in vertebrate prospective floor plate, EvoDevo, 2018.
Lopez-Sanchez, C., Garcia-Martinez, V., Schoenwolf, G.C., 2001. Localization of cells of the prospective neural plate, heart and somites within the primitive streak and epiblast of avian embryos at intermediate primitive-streak stages. Cells, tissues, organs 169, 334-346.
Ribes V, Briscoe J. 2009, Establishing and interpreting graded Sonic Hedgehog signaling during vertebrate neural tube patterning: the role of negative feedback. Cold Spring Harb Perspect Biol., 1(2):a002014.
Новость подготовил © 09.02.2018 Станислав Кремнёв