
Область научных интересов: EvoDevo, эмбриогенез беспозвоночных животных, формирование и эволюция планов строения Metazoa.
Формирование эволюционных новшеств – один из фундаментальных вопросов биологии развития. Существуют различные определения для понятия «новшество». Самое универсальное было предложено Эрнстом Майром: эволюционное новшество - это новая структура, которая может выполнять новые функции. Классическими примерами эволюционных новшеств являются зуб нарвала, преобразованный в «рог», а также рисунок на крыльях бабочек. Для многих новшеств неизвестны гомологичные структуры, из которых они эволюционировали. В статье, опубликованной международным коллективом под руководством профессора Beverley Glover (University of Cambridge), рассматривается , как с помощью коопции (вовлечения) генетических регуляторных сетей (ГРС) в развитие негомологичных структур, у маргариток получились цветки-обманщики. Это новшество позволило цветкам имитировать самку насекомого, привлекать самцов- опылителей и, таким образом, экономить на выработке нектара.
Секрет успеха Gorteria diffusa кроется в темных пятнах, расположенных на цветках, образующих соцветие-корзинку (рис. 1). Исходно эти пятна, формирующие кольцо в центре соцветия, призваны указывать опылителям расположение нектара. Однако некоторые представители G. diffusa модифицировали этот признак и стали использовать его для обмана самцов-опылителей, стимулируя у них псевдокопуляцию. Оранжевые цветки с темными пятнами имитируют самок мух-жужжал Megapalpus capensis (семейство Bombyliidae) – основных опылителей этого растения. Авторами статьи были выделены 4 основных фенотипа соцветий, которые с разной степенью успеха обманывали самцов-опылителей (рис. 1A). У фенотипа Naries темные пятна образуют кольцо в центре соцветия, и поверхность пятен гладкая. Такие соцветия не обманывают опылителей. Второй фенотип, Garies, имеет больший успех среди самцов-опылителей. У него пятна также образуют кольцо, однако на их поверхности присутствуют многоклеточные выросты -- папиллы (рис. 1B). Фенотип Okiep еще лучше имитирует сидящих на соцветии самок мух: пятна на лепестках хоть и гладкие, но не образуют замкнутого кольца. Успешнее всех обманывают опылителей представители четвертого фенотипа – Springbok. Они сочетают в себе лучшие качества Garies и Okiep, а именно дискретное расположение темных пятен и папиллы на их поверхности. Таким образом, в сложном эволюционном новшестве, коим являются цветки-обманщики, авторам удалось выявить основные морфологические модули: (1) темный цвет пятен, (2) структура поверхности пятен, (3) взаимное расположение пятен. Затем они последовательно разобрали молекулярные механизмы формирования каждого из этих модулей.
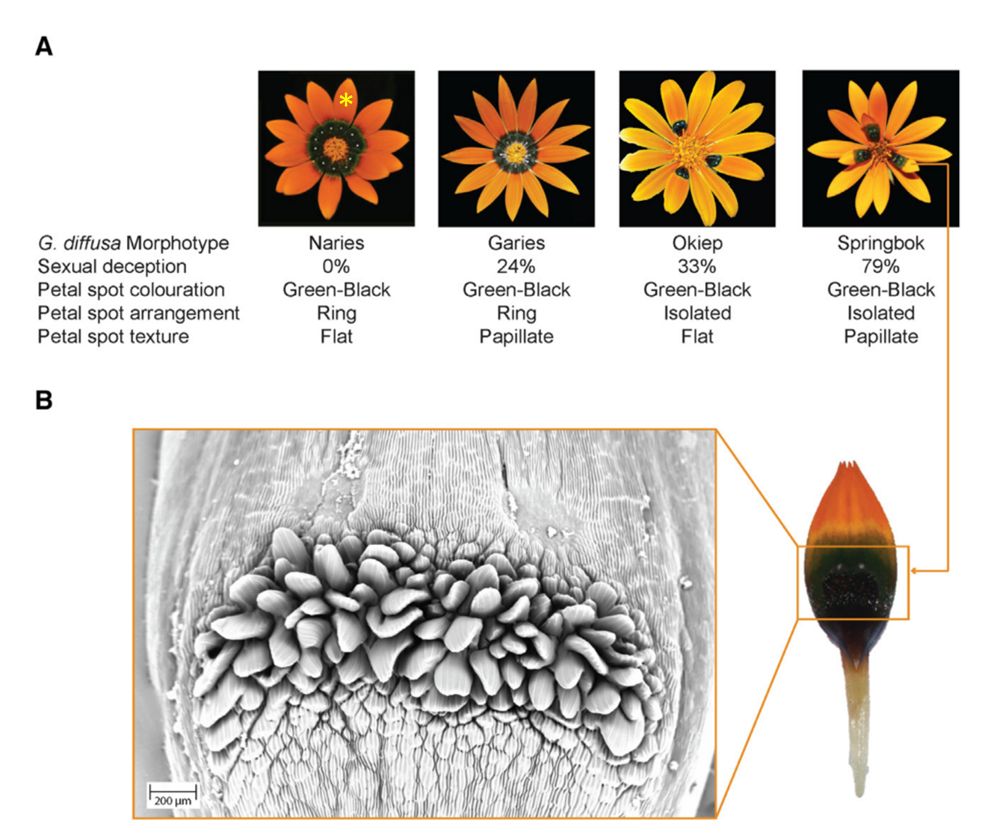
Рис. 1. Внешний вид соцветий Gorteria diffusa. A – Морфотипы соцветий G. diffusa. Желтой звездочкой обозначен отдельный цветок в составе соцветия-корзинки. B – Изображение поверхности темного пятна на цветке наиболее обманчивого морфотипа Springbok, полученное методом криоэлектронной микроскопии. Видно, что поверхность пятна у основания цветка покрыта папиллами (из Kellenberger et al., 2023).
Формирование темных пятен на лепестках связано с преобразованиями пигментов- антоцианов: цианидин-3-глюкозида и малонилированного цианидина. Эти пигменты экспортируются в вакуоли темных клеток основания лепестка, где образуют комплекс с ионами железа (рис. 2). В результате цвет этих пигментов меняется с красного на темно- синий. Ионы железа транспортируются в клетки основания цветка, где расположено пятно, из более светлых клеток кончика цветка. Авторы выяснили, что активное накопление железа в вакуолях клеток пятна обусловлено коопцией генов, регулирующих гомеостаз железа. Эти гены относятся к ГРС, регулирующей биосинтез флавоноидов и антоцианов. Именно колокализация темно-синего хелатированного антоциана с оранжевыми каротиноидами создает характерную зелено-черную окраску пятен на цветках.
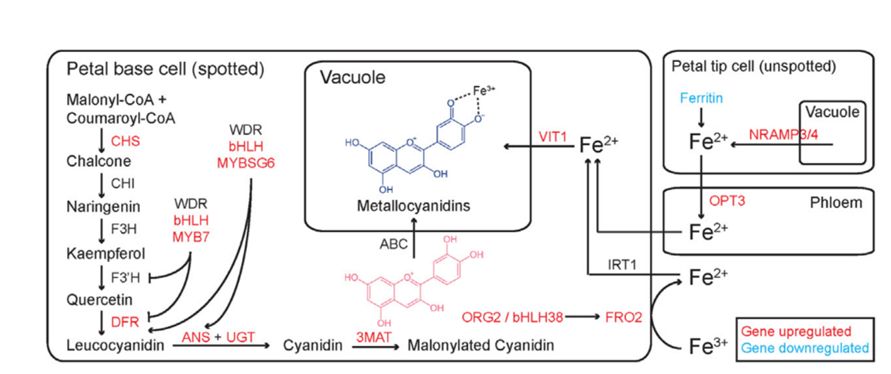
Рис. 2. Схема синтеза пигментов в клетках основания цветка фенотипа Springbok. Изменение цвета пигментов в клетках основания цветка происходит за счет формирования комплексов с ионами Fe2+ , которые импортируются из клеток кончика цветка (из Kellenberger et al., 2023).
Механизм, обеспечивающий развитие папилл в основании лепестка, оказался еще более необычным. В клетках, формирующих папиллы, активно экспрессируется ген GdEXPA7, который регулирует развитие корневых волосков у растений. Продукт экспрессии этого гена разрыхляет клеточную стенку путем диссоциации составляющих ее гликанов. В результате клетки корневых волосков образуют характерные выросты. Как выяснилось, этот же механизм обеспечивает формирование папилл на цветках G. diffusa.
Наибольшим «обманным потенциалом» обладают соцветия морфотипа Springbok, у которого пятна расположены не на всех цветках, а только на некоторых из них. Это позволяет имитировать сидящих на соцветии самок мух. У маргариток самые первые цветки формируются ближе к центру соцветия. И именно на них образуются темные пятна. Более молодые цветки закладываются между старыми по спирали и пятен уже не несут (рис. 3). Авторы предполагают, что изменение паттерна расположения пятен обусловлено вовлечением в регуляцию развития соцветия генов GdSPL1 и GdmiR156, которые у других растений регулируют переход между стадиями жизненного цикла (вегетативной и генеративной фазами). Развитие первых (пятнистых) цветков у G. diffusa сопровождается экспрессией гена GdSPL1. При формировании более молодых цветков экспрессия GdSPL1 подавляется продуктом гена GdmiR156, и поэтому на более молодых цветках пятна не формируются.
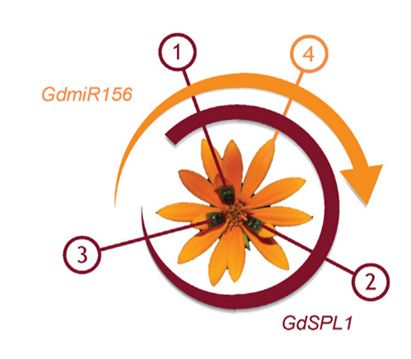
Рис. 3. Модель развития соцветия морфотипа Springbok. Формирование цветков происходит по кругу под углом 132 o . В ходе развития первых нескольких цветков (в данном случае, цветков 1 – 3) активен ген GdSPL1, который инициирует развитие пятен. При развитии более поздних цветков (4 и далее), активируется ген GdmiR156, который подавляет экспрессию GdSPL1 (из Kellenberger et al., 2023).
О том, что коопция генов и модулей ГРС сопряжена с формированием эволюционных новшеств, известно давно. Тем не менее, эта статья примечательна подходом авторов к исследованию данной проблемы. Во-первых, в данной работе проблема эволюционных новшеств изучается на растительном объекте, что пока ещё редкость. Во-вторых, авторы смогли выделить в сложном морфологическом новшестве независимые модули, что позволило подробно изучить механизм формирования каждого из них на молекулярно- генетическом уровне. Наконец, это одна из немногих статей, в которых описана четкая взаимосвязь между степенью эволюционного развития новшества и его адаптивной ценностью.
Новость подготовила
© Е.И. Андронова
08.04.2024