 © Дьяконова Варвара Евгеньевна,
© Дьяконова Варвара Евгеньевна,
доктор биологических наук, профессор РАН. Ведущий научный сотрудник лаборатории нейробиологии развития.
Область научных интересов: нейроэтология, нейробиология, нейротрансмиттеры, поведение.
Появление новых методов позволяет решать не только новые задачи, но иногда помогает ответить на старые классические вопросы, оставшиеся в свое время без ответа из-за отсутствия адекватных по трудоемкости подходов. Хорошим примером возвращения к старым вопросам на новом уровне является работа, опубликованная журналом Frontiers in Molecular Neuroscience в феврале 2021 года. Статья полностью лишена экспериментальной составляющей, она выполнена на основе данных, опубликованных другими исследователями.
Быстро набирающее скорость секвенирование транскриптомов одиночных клеток привело к тому, что в общем доступе находятся понейронно отсеквенированные нервные системы уже многих видов животных. Этими данными воспользовались швейцарские исследователи, чтобы окончательно разобраться с тем, как часто в нервной системе наблюдается котрансмиссия (использование одним нейроном нескольких нейротрансмиттеров) на широкой эволюционной шкале от кишечнополостных (гидра) до млекопитающих (мышь). Кроме того, на некоторых видах им удалось проверить, может ли меняться число нейронов с колокализацией нейротрансмиттеров в онтогенезе. Были использованы данные по транскриптомам нейронов гидры (Hydra vulgaris), головного ганглия планарии (Schmidtea mediterranea), нервной системы нематоды (Caenorhabditis elegance), головного мозга личинки и взрослой мухи (Drosophila melanogaster), головного мозга сидящего позвоночного асцидии (Ciona intestinalis), рыбы (Danio rerio), конечного мозга рептилий черепахи и ящерицы (Trachemys scripta и Pogona vitticeps) и, наконец, мозга молодых и старых мышей (Mus musculus). Изучали случаи колокализации только классических нейротрансмиттеров (глутамат, ацетилхолин, ГАМК, глицин, моноамины), используя наличие в транскриптомах нейронов маркерных РНК (отвечающих за синтез и везикулярный перенос соответствующих сигнальных молекул).
Часто колокализацию нейротрансмиттеров рассматривают как нарушение принципа Дейла, сформулированного в пятидесятые годы прошлого века и имевшее значение, сходное по значимости с выводом Рамона Кахаля о том, нервная система - это не синцитий, а совокупность отдельных нервных клеток. Принцип Дейла усиливал представление о независимой сущности нейрона. Однако у самого принципа Дейла существует два разных прочтения. Упрощенное предполагает, что один нейрон может использовать только один нейротрансмиттер, который секретируется всеми его окончаниями. Часть исследователей интерпретирует его иначе, как постоянство набора нейротрансмиттеров для всех секретируемых областей одного нейрона. О том, что принцип Дейла в упрощенном варианте нарушается, было известно давно. Существуют примеры колокализации классических нейротрансмиттеров с нейропептидами, газообразными сигнальными молекулами и у первично-, и у вторичноротых животных. Функциональное значение такой колокализации также изучалось и обсуждалось. Но вот, насколько это явление распространено в отношении классических нейротрансмиттеров оставалось не совсем ясным. Котрансмиссия– это экзотика или правило для нервной системы?
Сторонники химической организации мозга рассматривали колокализацию нейротрансмиттеров скорее как исключение из правила. Предполагалось, что она нужна в редких случаях для специальных целей, либо отражает случайное «недоразвитие» трансмиттерной специфичности. Эта точка зрения вполне понятна, если исходить из того, что раные нейротрансмиттеры имеют разные функции в нервной системе. В этом случае колокализация не может быть правилом для нервной системы. Для того, чтобы подтвердить или опровергнуть эти представления, 10-15 лет назад потребовалась бы работа нескольких институтов. Ведь, ранее единственный метод, который позволял выявлять колокализацию нейротрансмиттеров, было иммуногистохимическое окрашивание, иногда в сочетании с гибридизацией in situ. Сейчас подробный анализ явления у далеких в систематическом отношении организмов выполнен всего двумя соавторами.
Итак, что же выяснилось благодаря клеточной траскриптомике? Перечислим коротко основные результаты этой работы.
- Большинство нейронов (> 70 %) у всех исследованных видов экспрессирует маркеры только к одному нейротрансмиттеру.
- Нейротрансмиттерный профиль нервной системы существенно отличается у разных видов (например, у гидры классические нейротрансмиттеры представлены ацетилхолином и ГАМК, у рептилий основной фенотип представлен глутаматэргическими нейронами и в меньшей степени ГАМК, у мышей это соотношение меняется на обратное: преобладание ГАМК над глутаматом, к тому же существенно возрастает доля моноаминэргических клеток).
- Колокализация классических нейротрансмиттеров неизменно выявляется у всех исследованных видов примерно в 10-25% нейронов (Рис. 2). При этом минорные нейротрансмиттерные фенотипы (более редкие) могут быть представлены нейронами, синтезирующими еще один или даже несколько классических нейротрансмиттеров в 50% случаев и больше.
- Зависимость частоты колокализации от эволюционной удаленности организмов не выявлена.
- В онтогенезе число мультитрансмиттерных нейронов может меняться. В нервной системе взрослых дрозофил значимо снижается доля мультитрансмитттерных клеток по сравнению с нервной системой личинки (Рис 3 верх). У старых мышей она существенно не меняется по сравнению с молодыми (Рис. 3 низ).
- Кластерный анализ не выделяет нейроны с колокализацией нейротрансмиттеров в отдельные группы, как правило, они входят в кластеры нейронов, секретирующих один из ко-трансмиттеров (Рис. 4, 5). С другой стороны, не у всех видов проявляется и четкая кластеризация монотрансмиттерных клеток.
Правило или исключение? Теперь, когда у нас есть развернутая картина, оказалось, что однозначно ответить на этот вопрос все-равно не получится. С одной стороны, большинство нейронов у всех исследованных видов использует только один классический нейротрансмиттер, а нейроны с ко-трансмиссией принадлежит к кластерам нейронов, вырабатывающих один из ко-трансмиттеров, что согласуется скорее со случайным, а не направленным происхождением этих клеток. Уменьшение доли колокализаций при переходе от личиночного к взрослому мозгу тоже вроде бы согласуется с представлениями о котрансмиссии классических медиаторов как о случайном недоразвитии трансмиттерной специфичности. С другой стороны, доля нейронов с колокализацией явна велика для случайной, а настойчивое проявление котрансмиссии в эволюции от гидры до млекопитающих также свидетельствует о том, что даже 30 % нейронов с обобщенным трансмиттерным фенотипом (личинка дрозофилы) не мешает работе нервной системы. Суха теория, мой друг, а древо жизни вечно зеленеет.
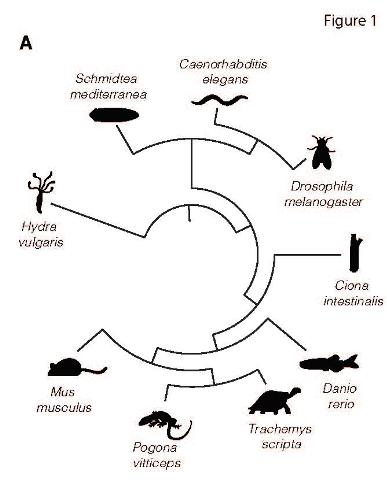
Рис.1. Исследованные виды животных на круговом филогенетическом дереве.
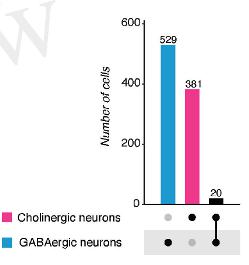
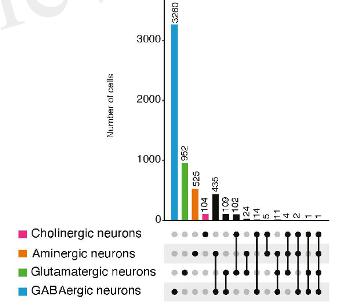
Рис.2. Соотношение трансмиттер-специфичных нейронов (цветные) и нейронов с колокализаций (черные) у гидры и мыши. Вертикальные линии указывают, какие именно нейротрансмиттеры колокализованы.
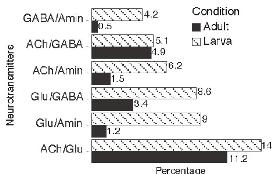
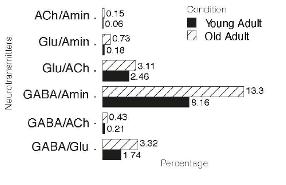
Рис.3. Изменения соотношения нейронов с колокализацией нейротрансмиттеров в онтогенезе (в процентах от общего числа нейронов). Верху личинки и взрослые организмы Drosophila. Внизу мозг молодых и старых мышей.

Рис.4. Нейроны, экспрессирующие везикулярные транспортеры к ацетилхолину (розовые) и глутамату (зеленые), на кластерных картах (UMAP plots). Белыми стрелками отмечены нейроны с ко-экспрессией обоих транспортеров. Видно, что в данном случае, они относятся к кластеру глутаматэргических нейронов.
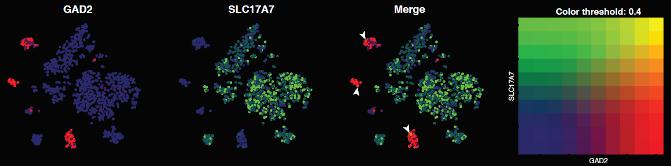
Рис.5. Нейроны, экспрессирующие фермент синтеза ГАМК GAD2 (розовые) и везикулярный транспортер глутамата 1 SLC17A7 (зеленые), на кластерных картах (UMAP plots). Белыми стрелками отмечены нейроны с ко-экспрессией обоих генов. Видно, что в данном случае, они все относятся к кластеру гамкэргических нейронов.
Новость подготовила © Дьяконова В.Е.
03.08.2021