 © Григорян Элеонора Норайровна. зав. лабораторией проблем регенерации. д.б.н.
Область научных интересов: клеточные и молекулярные механизмы регенерации органов и тканей у позвоночных животных.
© Григорян Элеонора Норайровна. зав. лабораторией проблем регенерации. д.б.н.
Область научных интересов: клеточные и молекулярные механизмы регенерации органов и тканей у позвоночных животных.
Хорошо известна уникальная способность саламандр регенерировать целые органы, включая сложно организованные, такие как конечность. Как и за счет чего происходит этот процесс? Какими должны быть условия у млекопитающих и человека для воспроизведения способности отращивать конечность de novo?
Со времени открытия эпиморфной регенерации конечности у низших позвоночных в XVIII веке ее изучение не прекращалось. В прошлом веке постепенно сложились представления о том, что для корректной регенерации конечности необходимы заживление раны, иннервация, формирование бластемы, последующий рост и дифференцировка бластемы. В 70-х годах (Bryant, 1976) стало понятно, что для осуществления регенерации конечности и формирования ее бластемы клетки антериальной и проксимальной областей конечности должны взаимодействовать (рисунок). При этом не было понятно, какого рода клеточные и молекулярные взаимодействия имеют место, какой именно «диалог» (antero-posterior crosstalk) происходит. И вот недавно, в журнале «Nature» опубликована статья, выполненная группой авторов из DFG исследовательского центра регенеративной терапии в Дрездене, проливающая свет на этот вопрос. В статье аргументируется необходимость присутствия и взаимодействия всего лишь двух факторов для осуществления полной регенерации конечности, а именно лигандов сигнальных путей Shh и Fgf8. Источником Shh оказались клетки постериальной области, источником Fgf8 – антериальной. Для понимания этого, казалось бы, простого факта потребовалось выстраивание достаточно хитроумной многошаговой экспериментальной работы. Были использованы хирургические манипуляции, трансплантации, иммуногистохимия, обработка химическими агонистами и ингибиторами, направленные инъекции бакуловирусов, несущих интересующие гены. Вначале, авторы использовали ранее полученные результаты, согласно которым при повреждении только антериальной области и подведении туда нерва образуется неразвивающаяся бластема (“frozen” blastema). Это позволило определить, чего не достает в молекулярном отношении для развития полноценного регенерата конечности. «Разморозить» ситуацию удалось при помощи «имитатора» активности сигнального белка Sonic hedgehog (Shh) - белка SAG (smoothened agonist). Его аппликация привела к формированию конечности на антериальной стороне конечности. Другими словами, была воспроизведена ситуация связывания рецепторами секретируемого клетками постериальной бластемы Shh лиганда, запускающая последующие клеточные регенеративные ответы. Активация hedgehog сигнального пути оказалась достаточной, чтобы активировать и завершить регенерацию в антериальной бластеме, но при поддержке экспрессии сигнального пути FGF – ключевого условия для отрастания бластемы. Ранее, было известно, что ни антериальные, ни постериальные бластемы не могут независимо осуществить полноценную регенерацию. Авторы обнаружили, что постериальная часть не может активировать и поддерживать устойчивую экспрессию FGF8 сигналинга в ответ на Shh. Экспериментальное усиление FGF8 экспрессии в постериальной области конечности приводило к росту дополнительной конечности только при условии, что Shh сигнальный путь по-прежнему активен, его ингибирование блокировало регенерацию. Вопрос, почему усиленная экспрессия FGF8 в отсутствие Shh сигналинга не может промотировать рост бластемы на постериальной стороне, остается пока без ответа.
Таким образом, эта работа показала, что у аксолотля антериальные и постериальные области конечности содержат клетки, ограниченные в своей компетенции комплементарно экспрессировать растворимые сигнальные факторы, необходимые для роста и правильного формирования вновь образующейся бластемы со стадии «средней почки». Эти данные лежат в русле ранее высказанной гипотезы о существовании границ взаимовлияний в тканях культи и регенерате конечности (Meinhardt, 1983), когда экспрессия диффундируемых сигнальных молекул индуцируется на границах больших топографических доменов конечности. Работа Nacu et al. (2016) несомненно, является шагом, приближающим нас к индукции регенерации конечности у млекопитающих с помощью набора определенных факторов. Однако при этом отметим, что исследование проведено на неотенической педоморфной форме Urodela и описанную стратегию клеточных взаимодействий при разработке подходов к восстановлению тканей конечности у взрослых тетрапод не следует полагать абсолютно универсальной.
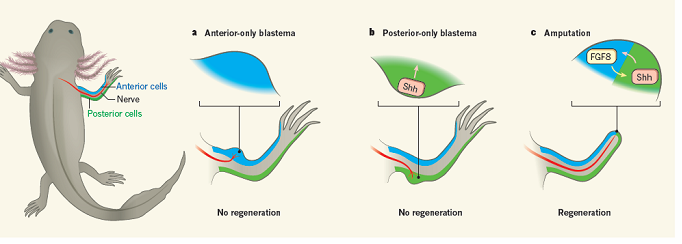
Рисунок. Механизм взаимодействия сигнальных молекул Shh и FGF8, необходимого для роста и дифференцировки бластемы при регенерации конечности у аксолотля. (а) – неразвивающаяся бластема с подведенным нервом на антериальной стороне передней конечности, (b) – неразвивающаяся бластема с подведенным нервом на постериальной стороне передней конечности, (с) – осуществление регенерации после ампутации при иннервации и взаимодействии Shh и FGF8 на границе, секретируемых клетками антериальной и постериальной областей.
Из: Torres, 2016
Copyright © 2016 Grigoryan E.N.