https://www.mdpi.com/2073-4425/11/4/386
 © Баклушинская Ирина Юрьевна, доктор биологических наук, в.н.с. лаборатории эволюции генома и механизмов видообразования.
© Баклушинская Ирина Юрьевна, доктор биологических наук, в.н.с. лаборатории эволюции генома и механизмов видообразования.
Область научных интересов: хромосомное видообразование, детерминация пола.
Число хромосом и особенности их формы - стабильная характеристика, специфичная для вида. Вместе с тем, даже для человека характерны многочисленные хромосомные мутации, среди них с наибольшей частотой (1 на 1000) обнаруживаются так называемые робертсоновские транслокации, полагают (King, 1993), что именно робертсоновские перестройки сыграли важнейшую роль в эволюции млекопитающих. Это особый тип перестроек, при котором происходит объединение двух одноплечих хромосом в одну, двуплечую. Некоторые исследователи считает, что это простое слияние, но другие противятся термину "fusion" - слияние. Дело в том, что у одноплечих (акроцентрических) хромосом есть второе, маленькое плечо, которое с большой долей вероятности теряется при такой мутации, а значит это сложный процесс, а не просто слияние. Также неясно, что происходит с такой загадочной структурой как центромера при объединении двух хромосом, возникают ли сначала дицентрики, а потом одна центромера становится неактивной, либо этот фрагмент хромосомы удаляется еще перед слиянием.
Существует несколько моделей, созданных для объяснения механизма формирования робертсоновских транслокаций. В самой ранней модели (Sip) ведущая роль отводится теломерам, роль которых трудно переоценить (Оловников, 1971). Более поздние работы учитывают пространственное расположение хромосом в ядре, прикрепление к ядерной оболочке, движение хромосом в ядре в ходе клеточного цикла и другие особенности (Cremer, Cremer, 2001; Solovei et al., 2009). Вместе с тем все современные модели ориентированы на решение проблемы возникновения транслокаций для интерфазного ядра и так или иначе сводятся к двум вопросам: что было вначале, разрыв ДНК или контакт хромосом ("breakage first" или "contact first")? (Farré et al., 2009). Взаимное расположение хромосом может облегчить или затруднить контакт, но вопрос о том, почему негомологичные хромосомы движутся навстречу друг другу и вступают в контакт, пока не изучен совершенно. Следует подчеркнуть, что эти исследования имеют не только академическую ценность, известны многочисленные нарушения хромосомного набора в канцерогенезе, специфичные для различных типов рака, вплоть до формирования специфических перестроек, делающих клетку устойчивой к химиотерапии. Вместе с тем, еще раз подчеркнем, что все вышесказанное относится к митотическим делениям, наиболее ярко такие нарушения проявляются в соматических тканях, наследоваться же перестройки могут в том случае, если они возникли в ходе созревания половых клеток, до стадии мейоза.
Объяснение наследования перестроек всегда являлось ахиллесовой пятой концепции хромосомного видообразования. Очевидно, что необходима модель формирования перестроек в мейозе, но это строго детерминированный процесс с многочисленными "проверочными пунктами" (check points), что делает почти невозможным получение измененных гамет. Хромосомы в мейозе изменяются по строго детерминированной программе, их передвижения иногда сравнивают с классическим балетом (Page, Hawley, 2003). Известен лишь один эволюционно значимый механизм, который на уровне мейоза может приводить к образованию новых видов с измененными хромосомными наборами - мейотический драйв (de Villena, Sapienza, 2001). Из-за неравнозначности женского мейоза возможно неравновесное расхождение хромосом в полярные тельца при двух последовательных делениях мейоза и, в итоге, соотношение яйцеклеток гетерозиготной матери может быть нарушено в пользу тех, что несут измененный кариотип. Но и в этом случае вопрос о возникновении самой перестройки остается открытым.
Многообразие живых существ и в данном случае оказалось источником новых знаний. Ряд видов млекопитающих демонстрирует значительную изменчивость хромосомных наборов из-за робертсоновских транслокаций, причины этого явления неясны. Среди наиболее известных - домовая мышь, обыкновенная бурозубка, туко-туко и слепушонки. Мы много лет изучали восточную слепушонку из Памиро-Алая, где был описан своеобразный взрыв хромосомной изменчивости (Lyapunova et al., 1980), но лишь недавно, благодаря использованию хромосомного пейнтинга удалось оценить масштабы этого явления (Romanenko et al., 2019). Также были на современном уровне изучены хромосомы алайской слепушонки (Bakloushinskaya et al., 2019). Этот зверек, живущий под землей (рис. 1), внешне не отличается от восточной слепушонки; но в 1969 г. был описан как новый вид на основании числа хромосом (52, а не 54, как у восточной), сниженного из-за появления гигантской пары двуплечих хромосом в наборе (Воронцов и др., 1969). Как и родственных видов, восточной и обыкновенной слепушонки, у алайской самцы утратили Y хромосому, в их хромосомном наборе, как и у самок, две X хромосомы. Система детерминации пола в этой группе млекопитающих до сих пор неизвестна (см. обзор Bakloushinskaya, Matveevsky, 2018).

Рис. 1. Алайская долина, алайская слепушонка
Оказалось, что хромосомные наборы алайской слепушонки изменяются чрезвычайно быстро, робертсоновская перестройка может закрепиться в течение всего 30 лет! Очевидно, должен быть какой-то механизм для столь быстрой эволюции хромосомного набора. И этот механизм был нами обнаружен при изучении мейоза у слепушонок из Алайской долины. Мы увидели, что танец хромосом в профазе мейоза I у алайской слепушонки имеет особенности, нехарактерные для других видов млекопитающих. Итак, что же мы увидели? Картины мейоза, которые нам показали хромосомы слепушонок, оказалось возможным разделить на четыре основные стадии, первые три из которых больше всего напоминали старинный танец - менуэт.
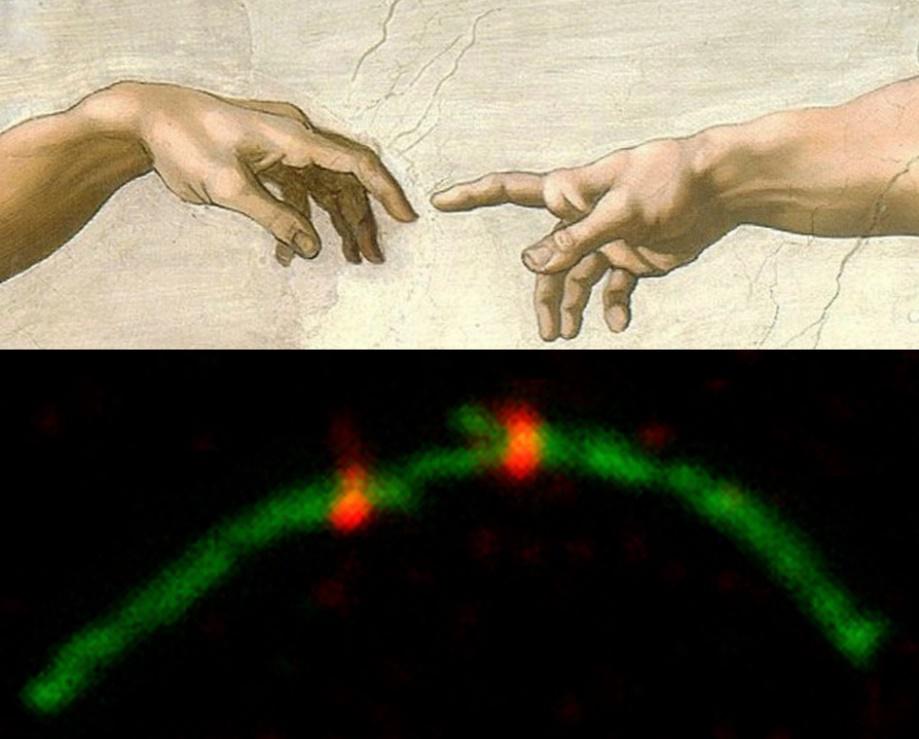
Первая и наиболее изменчивая стадия - сближение. [Мы хотели назвать эту стадию "намерение" (intention), однако научные редакторы указали нам на антропоморфный оттенок этого термина]. Хромосомы подходят ближе друг к другу, маленькие плечи удлиняются и становятся похожи на грациозно протянутую руку. И затем, маленькими шажками (как танцоры в менуэте) хромосомы приближаются друг к другу (рис. 2). Сходная картина известна для мейоза некоторых форм домовой мыши, акроцентрические хромосомы могут собираться парами или группами вокруг облака гетерохроматина в течение ранней пахитены.

Огюст Бувье (Augustus Bouvier) Менуэт 1874 |
Хромосомы: сближение, пахитена |
Рис. 2. Сближение.
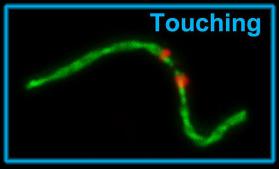
Во время второй стадии хромосомы слепушонок прикасаются друг к другу этими вытянувшимися плечами - "прикосновение", рис. 3. Эта и последующие стадии ранее не были описаны для каких-либо других видов живых существ.

Иоганн Цоффани (Johann Zoffany) Семейный вечер: менуэт, 1780-178 |
Хромосомы, прикосновение |
Рис. 3. Прикосновение.
Третья стадия - упрочение контакта, партнеры оказываются еще ближе друг к другу - "связывание" (рис. 4).

Дункан Маккелар (Duncan Mackellar), 1908, менуэт, фрагмент |

Хромосомы, связывание |
Рис. 4. Связывание.
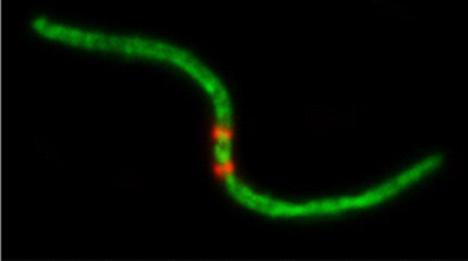
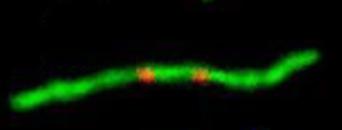
И, заключительная, самая редкая, но и наиболее важная стадия - "слияние" (рис. 5). Мы видели такие слившиеся хромосомы с двумя центромерами также и у восточной слепушонки.
a) |
b) |
Рис. 5. Слияние. Двуплечие хромосомы с двумя центромерными сигналами: а) алайская слепушонка, б) восточная слепушонка.
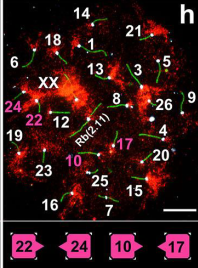
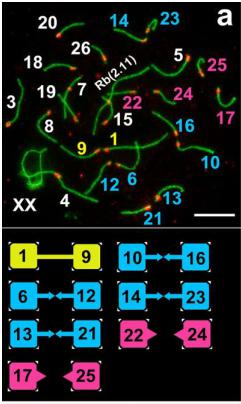
Удивительно, но нам не удалось выявить четкой закономерности для контактов хромосом, они все время пытались составить разные пары! Это, впрочем, характерно и для менуэта (рис. 6). Лидером по числу контактов оказалась хромосома №1, самая большая по размеру, которая была нами замечена ранее в транслокации у слепушонок из популяции, обитающей за сотни километров от Алайской долины.

Фредерик Каммерер, менуэт (Frederik Hendrik Kaemmerer) |
Хромосомы, пахитена. |
Рис. 6. В поисках партнера.
Итак, мы предполагаем, что если не единственным, то одним из механизмов формирования робертсоновских транслокаций может быть изменение поведения хромосом в профазе мейоза I. Формирование контактов негомологичных хромосом вплоть до слияния может привести к наследованию такого рода перестроек. На основе полученных результатов мы предложили новую модель формирования робертсоновских транслокаций, в которой первым шагом является контакт хромосом в мейозе: "contact first in meiosis".
Воронцов Н.Н., Ляпунова Е.А., Закарян Г.Г., Иванов В.Г. Кариология и систематика рода Ellobius (Microtinae, Rodentia) // В сб. Мат-лы ко II Всесоюзному Совещанию по млекопитающим. Млекопитающие, эволюция, кариология, систематика, фаунистика. 1969. Новосибирск. С. 127-129.
Оловников А.М. Принцип маргинотомии в матричном синтезе полинуклеотидов // ДАН. 1971. Т. 201. N 6. С. 1496-1499.
Bakloushinskaya I., Lyapunova E.A., Saidov A.S., et al. Rapid chromosomal evolution in enigmatic mammal with XX in both sexes, the Alay mole vole Ellobius alaicus Vorontsov et al., 1969 (Mammalia, Rodentia). Comp. Cytogenet. V. 2019. V. 13(2). P. 147-177. https://compcytogen.pensoft.net/article/34224/
Bakloushinskaya I., Matveevsky S. Unusual ways to lose a Y chromosome and survive with changed autosomes: a story of mole voles Ellobius (Mammalia, Rodentia) // OBM Genetics. 2018. V.2 (3). http://www.lidsen.com/journals/genetics/genetics-02-03-023
Cremer T., Cremer C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells// Nat. Rev. Genet. 2001. V. 2(4). P. 292-301.
de Villena F.P., Sapienza C. Female meiosis drives karyotypic evolution in mammals. Genetics // 2001. V. 159. P. 1179-1189.
Farré M., Robinson T.J., Ruiz‐Herrera A. An Integrative Breakage Model of genome architecture, reshuffling and evolution: The Integrative Breakage Model of genome evolution, a novel multidisciplinary hypothesis for the study of genome plasticity // BioEssays. 2015. V. 37(5). P. 479-488.
Lyapunova E.A., Vorontsov N.N., Korobitsina K.V. et al. A Robertsonian fan in Ellobius talpinus // Genetica (Hague). 1980. V. 52/53. P. 239–247.
Page S.L., Hawley R.S. Chromosome choreography: the meiotic ballet // Science. 2003. V. 301(5634). P.785-789.
Romanenko S.A., Lyapunova E.A., Saidov A.S. et al. Chromosome translocations as a driver of diversification in mole voles Ellobius (Rodentia, Mammalia) // Int. J. Mol. Sci. 2019. V. 20(18). 4466. https://www.mdpi.com/1422-0067/20/18/4466
Solovei I., Kreysing M., Lanctôt C., et al. Nuclear architecture of rod photoreceptor cells adapts to vision in mammalian evolution // Cell. 2009. V. 137(2). P. 356-368.
Новость подготовила © Баклушинская И.Ю.
04.05.2020