https://doi.org/10.1038/s41557-022-01111-y
 © Баклушинская Ирина Юрьевна, доктор биологических наук, главный научный сотрудник лаборатории эволюции генома и механизмом видообразования ИБР РАН.
© Баклушинская Ирина Юрьевна, доктор биологических наук, главный научный сотрудник лаборатории эволюции генома и механизмом видообразования ИБР РАН.
Недавняя статья М. Никитина вызвала довольно бурное обсуждение в сети, причем большая часть комментариев молекулярных биологов оказалась негативной. Практически все исследователи, рассуждая о "новизне и фундаментальной значимости" предложенной концепции, приходят к выводу, что нет ни того, ни другого. Для корректности оценки предлагаю прочитать исходную статью, здесь я лишь кратко изложу собственные соображения.
М. Никитин описывает феномен «коммутации цепей» —механизм хранения и обработки информации ДНК/РНК, основанный на обратимых низкоаффинных взаимодействиях некомплементарных нуклеиновых кислот. Варианты этих взаимодействий определяются «сродством» молекул и в общем случае описываются открытым еще в XIX веке законом о зависимости скорости реакции от концентрации участвующих веществ. Для биологов новый взгляд, несомненно, интересен, давно высказывается предположение, что наряду с эпигенетическими механизмами, которые интенсивно изучаются последние десятилетия, существуют и другие механизмы хранения/изменения/передачи генетической информации.
Термина "commutation" нет в генетических словарях. Кембриджский словарь дает значения, используемое в финансовой сфере или юриспруденции, что мало помогает понять смысл в приложении к ДНК. В Meriam-Webster из большого количества значений и словоформ можно выбрать глагол commutate – происходящий от латинского commutare, в приложении к нашей теме - имеющий способность изменяться или уже изменённый. На мой взгляд, у русскоязычного читателя, скорее, возникает образ коммутатора – переключателя, например, телефонных звонков, которые были обычны в досотовую (аналоговую) эпоху телефонной связи. В новой концепции информация (=сигнал) передается благодаря взаимодействию молекул разной степени сродства. Максимальная аффинность достигается в случае полной комплементарность двух олигонуклеотидов. При постепенной замене оснований в одном из олигонуклеотидов аффинность будет уменьшаться, но при этом может быть сформирован "континуум аффинностей", который и обеспечивает коммутацию.
В статье приведена довольно наглядная схема к этому тезису (рис. 1).
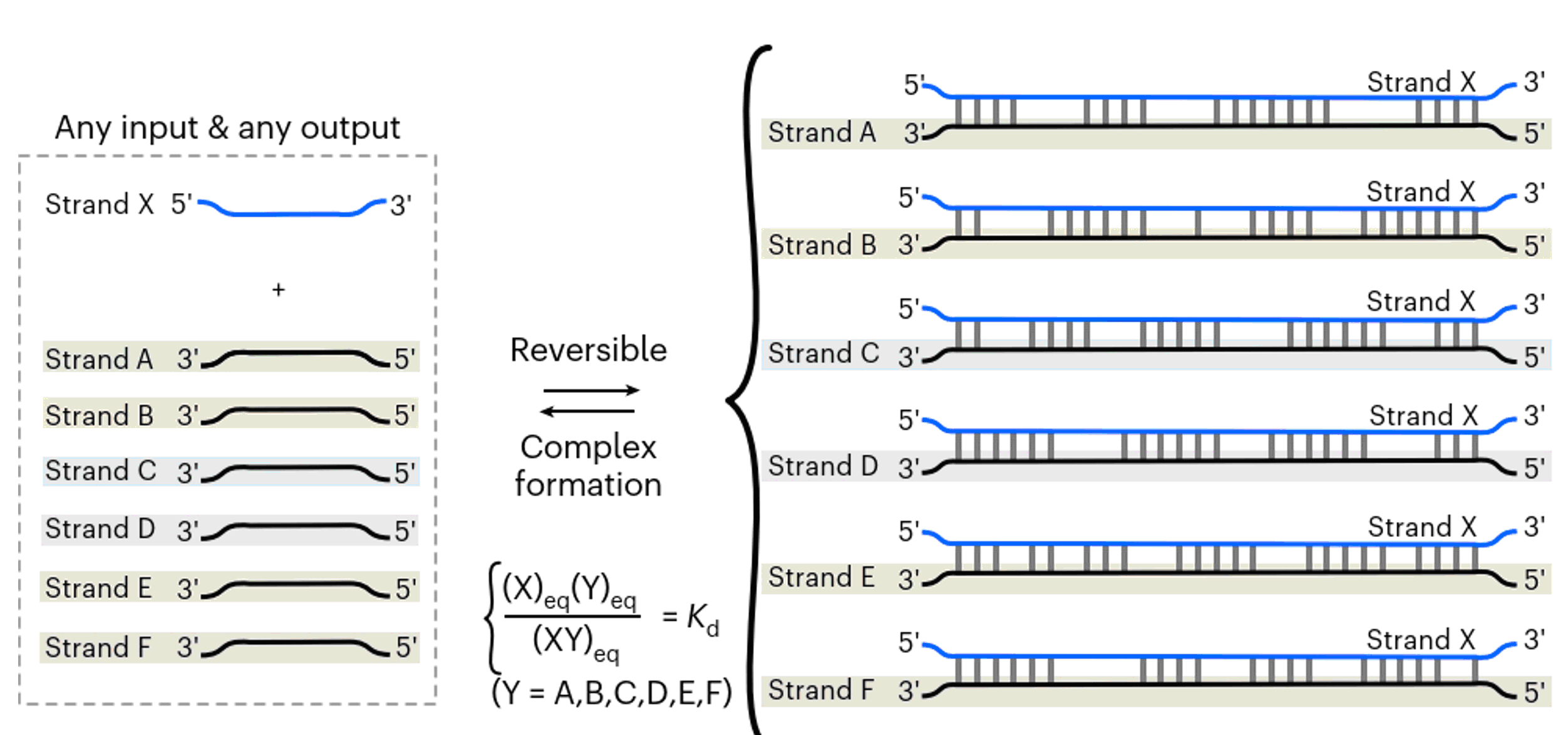
В качестве одного из примеров различий между классической парадигмой и новой концепцией, М. Никитин сравнивает количество вариантов олигонуклеотидов, которые могут как-то взаимодействовать с изучаемым фрагментом такого же размера. Так вот, фрагмент из 20 нуклеотидов предоставляет 2572 ≈ 10172 вариантов для воздействия на ген, в отличие от 420 ≈ 1012 в классической парадигме. И такое разнообразие возможностей обеспечивает произвольным неструктурированным коротким одноцепочечным ДНК специфичность регуляции гена, для чего обязательным является наличие в среде или организме других некомплементарных олигонуклеотидов. Т.е. триггер оказывается также неспецифичным. Возникает вопрос о надежности кажущегося хаотичным и ведущим к непредсказуемым последствиям способа передачи информации. На мой взгляд, именно тезис об обязательном наличии еще каких-то некомплементарных нуклеотидов является слабым звеном концепции. Если мы включаем систему неспецифичных матриц (матриц в понимании Н.К. Кольцова "Omnis molecula ex molecula" - "Каждая молекула от молекулы") в систему генетической регуляции, то получаем неконтролируемый уровень фенотипической изменчивости. Естественный отбор никто не отменял, именно он определит, что будет наследоваться. Вопрос в том, насколько расточительна или, напротив, экономна такая система функционирования генома.
Вероятно, можно поспорить с М. Никитиным в том, что никто не замечал и не учитывал в своей работе значения неполной комплементарности, например, праймеров, к тем последовательностям, которые нужно проанализировать. Более того, по его мнению, короткие фрагменты (например, 7-10 нуклеотидов), которые присутствуют практически в любых экспериментах, обычно рассматриваются как не заслуживающий внимания мусор. Здесь просматривается тонкий момент анализа результата эксперимента. Как правило, мы, сталкиваясь с тем, что результат эксперимента резко отличается от ожидаемого, готовы считать опыт неудачным и искать пути "исправления" для того, чтобы получить ожидаемое. А М. Никитин пошел по другому пути – предположил, что в генетических процессах неполное соответствие является закономерным и служит запрограммированным механизмом передачи информации. И в этом смысле посыл Уильяма Бейтсона о ценности исключений (Treasure your exceptions! W. Bateson) представляется особенно значимым.
В отличие от классической парадигмы, в рамках которой рибонуклеиновые кислоты стали центральным звеном наследственности (Watson, Crick, 1953) и точность передачи информации обеспечивает фундаментальную основу жизни, в предложенной М. Никитиным концепции низкоспецифичные молекулярные взаимодействия определяют способ обработки и хранения данных, позволяя более гибко и чутко обеспечивать настройку/регуляцию.
М. Никитин предлагает рассмотреть три аспекта применимости феномена некомплементарной коммутации: конструирование биокомпьютеров, биомедицина и природные эволюционные процессы. Нужно отметить, что М. Никитин подчеркивает, что явление молекулярной коммутации in vivo вряд будет возможно напрямую увидеть и доказать, хотя и существует вероятность случайной манифестации явления. В статье он описывает этот феномен двумя путями – предлагая математическую модель и проводя эксперименты in vitro с короткими олигонуклеотидами. Для оценки биологической значимости концепции, несомненно, интереснее эволюционный аспект. И в этом плане привлекает внимание ремарка о том, что наши знания о роли малых ДНК и РНК как, впрочем, и длинных некодирующих РНК, крайне недостаточны (см. также обзор Mattick, 2023), а данная концепция может оказаться шагом к разработке проблемы. И, что так же интересно, привлекает внимание к другим молекулам – протеинам, малым молекулам, липидам. А мы помним, что Н.К. Кольцов сформулировал матричный принцип для полимеров, а именно, белковых молекул.
Насколько будет востребована представленная М. Никитиным концепция коммутации, покажет время. Маловероятно, что эти новые представления будут быстро восприняты сообществом. Если в прошлом веке идея о непостоянстве генома представлялась крамольной, то сейчас без изучения «мусорной» ДНК, мобильных элементов, эпигенетических механизмов невозможно современную биологию (Хесин, 1984; Голубовский, 2000). Тем не менее, следует отметить, что несовершенство наших знаний о том, как функционирует геном, требует развития теоретических представлений. Мне хотелось бы выделить концепции А.М. Оловникова о парагеноме и экокроссинговере и концепцию Дж. Шапиро о способности генома к саморедактированию.
Концепции, предложенные Алексеем Матвеевичем Оловниковым, не раз существенно меняли мировоззрение биологов. На фоне его знаменитой работы о роли теломер (Оловников, 1973), другие статьи, возможно, меньше обсуждаются (Оловников, 2007, Olovnikov, 2009, 2022), но, вне всяких сомнений, имеют огромную ценность. В статье "Роль парагенома в развитии организмов", опубликованной в журнале Онтогенез в 2007 г., А.М. Оловников пишет, что «...текущие представления о геноме как единственной информотеке и единственной оперативной памяти организма принципиально неполны – не хватает одного важнейшего элемента, который можно обозначить как парагеном» ( с. 137). Парагеном — это "транзиторный набор коротких молекул ДНК, появляющихся на поверхности хромосом в ходе индивидуального развития для управления геномом". Парагеном состоит из трех групп носителей информации: принтомеры, хрономеры и филомеры. Название последней группы подчеркивает значение для эволюционных изменений, в данном случае, через модификации процессов развития. Статья крайне мало обсуждалась и цитировалась, что совершенно несправедливо, такой взгляд на функцию и эволюцию генома заслуживает повышенного внимания. Концепция экокроссинговера, о котором Алексей Матвеевич писал в 2022, через 15 лет после публикации статьи по парагеному, существенно дополняет ту самую «информотеку» в части возникновения неслучайной и адаптивной генетической изменчивости, предлагая рассмотреть циклические РНК в качестве инструмента для управления кроссинговером (Olovnikov, 2022, см. новость на сайте от 27.06.2022).
Американского микробиолог Дж. Шапиро высказал гипотезу о "самозаписывающем информацию геноме" ("The Read–Write (RW) genome") и развивает ее на протяжении более 20 лет (Shapiro, 2002, 2013, 2016, 2022). По его мнению, жизнь изменяет себя сама, используя природную генетическую инженерию, изменение генома может быть не случайным, а все происходящее находится под контролем естественного отбора. Эта концепция 10 лет назад была многими, в том числе М.Д. Франком-Каменецким (Frank-Kamenetskii, 2013), оценена как провокационная. На мой взгляд, эта гипотеза заслуживает внимания в первую очередь потому, Дж. Шапиро рассматривает геном (а не его фенотипическое осуществление) как предмет отбора, то есть естественный отбора (движущий или стабилизирующий) действует на уровне структуры нуклеиновых кислот, их упаковки, вплоть до хромосомного набора.
В этой связи концепция М. Никитина о роли аффинных взаимодействий и принципе коммутации информационного сигнала может рассматриваться в качестве нового направления в эволюционной биологии.
Список цитируемой литературы
Голубовский М.Д., 2000. Век генетики: эволюция идей и понятий. Научно-исторические очерки. СПб.: Борей Арт. 262 с.
Оловников А.М., 1971. Принцип маргинотомии в матричном синтезе полинуклеотидов. Доклады АН СССР. Т. 201. С. 1496–1499.
Оловников А.М., 2007. Роль парагенома в развитии организмов. Онтогенез. Т. 38(2). С. 136-158. / Olovnikov A.M., 2007. Role of paragenome in development. Russian Journal of Developmental Biology. V. 38(2). P. 104-123.
Хесин Р.Б. Непостоянство генома. М.: Наука, 1984. 472 с.
Bateson W.б 1908. The Methods and Scope of Genetics. Cambridge University Press. :“If I may throw out a word of counsel to beginners it is: Treasure your exceptions!… Exceptions are like the rough brickwork of a growing building which tells that there is more to come and shows where the next construction is to be.”
Frank-Kamenetskii M.D., 2013. Are there any laws in biology? Comment on “how life changes itself: The read–write (RW) genome” by James Shapiro. Physics of Life Reviews, 10(3), pp.328-330.
Mattick J.S., 2023. RNA out of the mist. Trends in Genetics, 39(3), pp.187-207.
Olovnikov A.M., 2009. Biological evolution based on nonrandom variability regulated by the organism. Biochemistry (Moscow), 74(12), pp.1404-1409.
Olovnikov A.M., 2022. Eco-crossover, or environmentally regulated crossing-over, and natural selection are two irreplaceable drivers of adaptive evolution: Eco-crossover hypothesis. Biosystems, 218, 104706.
Shapiro J.A., 2002. Genome organization and reorganization in evolution: formatting for computation and function. Annals of the New York Academy of Sciences, 981(1), pp.111-134.
Shapiro J.A., 2013. How life changes itself: The Read–Write (RW) genome. Physics of Life Reviews, 10(3), 287-323.
Shapiro J.A., 2016. Nothing in evolution makes sense except in the light of genomics: read–write genome evolution as an active biological process. Biology, 5(2), p.27.
Shapiro J.A., 2022. What we have learned about evolutionary genome change in the past 7 decades. Biosystems, 215, 104669.
Watson J. D., Crick, F.H.C., 1953. Molecular structure of nucleic acids: a structure for deoxyribose nucleic acid. Nature 171, 737–738.
Новость подготовила
© Баклушинская И.Ю.
03.04.2023