
Научные интересы: апиология.

Способность запоминать, в какое время и где можно найти пищу, дает фуражирующим насекомым важное эволюционное преимущество. Особенно сложные механизмы временной памяти развиты у социальных насекомых, таких как медоносные пчелы (Apis mellifera), которым нужны эффективные способы добычи пищи для поддержания колонии. Пчелы идеально подходят для изучения временной памяти, так как сочетают способность к запоминанию времени с относительно простой нейронной сетью циркадных часов и меньшим количеством генов по сравнению с млекопитающими (белые мыши) (Roy et al., 2025).
Временная память играет важную роль в фуражировочном поведении социальных насекомых, позволяя им связывать определенные события с конкретным временем и местом. Этот процесс называется пространственно-временным обучением (time-place learning, TPL). TPL основывается на временной памяти, которая управляется внутренним механизмом отсчета времени, известным как циркадные часы. Этот универсальный механизм помогает животным синхронизировать свою активность и физиологические процессы с определенным временем суток (Roy et al., 2025).
Первые исследования формирования временной памяти провел Белинг (Beling, 1929). Он обучил пчел посещать кормушку в фиксированное время дня и обнаружил, что они возвращались к пустой кормушке в последующие дни в то же время (Beling, 1929). Пчел можно было научить добывать пищу в любое время суток или даже в несколько временных интервалов в течение одного дня. Хотя Белинг (1929) и, продолживший его исследования Валь (Wahl, 1932), не использовали термин «циркадные часы», они описали основные характеристики циркадных ритмов в поведении пчел: 1) ритм у пчел работает автономно («free-run») с периодом около 24 часов без внешних временных сигналов, таких как циклы света и темноты или изменения доступности пищи (Zeitgebers); 2) ритмы у пчел могут синхронизироваться с окружающей средой через внешние сигналы, но только в определенных пределах, например, пчел нельзя обучить интервалам в 19 или 48 часов; 3) ритмы у пчел устойчивы к изменениям температуры, то есть их период остается неизменным при разных температурах (Beling, 1929; Wahl, 1932; Roy et al., 2025). Эти открытия показали, как временная память помогает пчелам оптимизировать фуражировочную деятельность.
Обучение месту и времени (time-place learning, TPL), описанное Белингом (Beling, 1929) и Валем (Wahl, 1932), требует от пчел умения одновременно ориентироваться во времени и пространстве (Beling, 1929; Wahl, 1932; Roy et al., 2025). Это более сложный процесс, чем запоминание отдельных сигналов, таких как запах или цвет, в разное время суток (Chouhan et al., 2015; Gonulkirmaz-Cancalar et al., 2023). Для TPL пчела должна объединять информацию о времени с набором признаков, таких как местоположение или другие параметры. Так, было показано, что пчелы могут связывать два запаха с двумя разными кормушками. Если добавить дополнительные сигналы, например цвет, пчелы успешно запоминают ассоциации с тремя местами кормления (Reinhard et al., 2006). Это говорит о том, что комбинация нескольких сигналов упрощает формирование временной памяти. Когда пчела воспринимает один из сигналов, например запах, она может вспомнить другие ассоциированные признаки, такие как цвет или маршрут к кормушке, что помогает ей вернуться к источнику пищи (Reinhard et al., 2004a, 2004b; Roy et al., 2025).
TPL обучение также обнаружено у позвоночных, таких как птицы (Biebach et al., 1989), мыши (Van der Zee et al., 2008) и рыбы (Reebs, 1996), а среди насекомых — в основном у перепончатокрылых, включая медоносных пчел (Beling, 1929; Wahl, 1932; Beer & Helfrich-Forster, 2020; Koltermann, 1971; Pahl et al., 2007), муравьев (Schatz et al., 1994; Harrison & Breed, 1987) и безжальных пчел (Murphy & Breed, 2008; de Jesus et al., 2014). Интересно, что TPL чаще встречается у эусоциальных насекомых, но пока неясно, является ли социальная организация важным фактором в формировании этого поведения. Исследования одиночных насекомых показали, что у двух видов одиночных пчел и бабочек Heliconiini также присутствовало TPL обучение (Toure et al., 2020; Gimenes et al., 1996). Предполагается, что развитие грибовидного тела — центров обучения и памяти у насекомых — может быть важным условием для формирования обучения TPL (Toure et al., 2020).
Одной из причин эволюции обучения TPL считают временную доступность цветочных ресурсов, которая меняется в течение дня в зависимости от вида растений (Bloch et al., 2017; Kleber, 1935). Насекомым выгодно фуражировать точно в период максимального нектаровыделения. Так, среди муравьев способности к обучению TPL проявлялись только у видов, питающихся нектаром (Schatz et al., 1994). В отличие от этого, плодовые мушки (Drosophila melanogaster), несмотря на развитую временную и пространственную память, не демонстрируют обучение TPL. Вероятно, это можно объяснить тем, что их питание фруктами не зависит от времени суток (Chouhan et al., 2015; Ofstad et al., 2011). Таким образом, способность запоминать только время может быть широко распространена среди насекомых (Chouhan et al., 2015), но сложные связи между временем и пространством, характерны только для видов, питающихся цветочным нектаром и пыльцой (Elizalde et al., 2020; Roy et al., 2025).
Начиная с работы Белинга (Beling, 1929), ученые изучали поведение пчел, обученных посещать кормушки в определенное время суток, как в естественных условиях (Beling, 1929; Wahl, 1932; Kleber, 1935; Beier & Lindauer, 1970; Frisch & Aschoff, 1987; Moore & Rankin, 1983; Moore et al., 2011; Renner, 1957, 1959), так и в контролируемых лабораторных садках с постоянными условиями (Wahl, 1932; Renner, 1957; Beier, 1968; Kefuss & Nye, 1970; Medugorac & Lindauer, 1967; Moore et al., 1989; Roy et al., 2025). Такие исследования помогли понять, как формируется пространственно-временная память у пчел.
В зависимости от времени, летную активность пчел можно разделить на четыре этапа: спад, предвкушение, пик активности и снижение (Roy et al., 2025). Было показано, что в день между тренировочными периодами пчелы вообще не летают к кормушке (спад). За несколько часов до ожидаемого времени кормления отдельные пчелы начинают приближаться к пустой кормушке, и их активность постепенно растет (предвкушение). Максимальная активность пчел у кормушки проявлялась во время тренировочного периода (пик активности) (Roy et al., 2025). После окончания тренировочного периода пчелы перестают посещать кормушку, и активность быстро падает (снижение). В тестовой ситуации, когда кормушка пуста, поведение пчел менялось: пчелы были наиболее активны в начале или немного ранее обученного времени кормления, но активность их резко падала, когда они не находили пищу (Moore et al., 1989). То, что пчелы прилетают чуть раньше, вероятно, имеет эволюционное преимущество: лучше прибыть заранее, чем опоздать. Особенности формирования временной памяти у пчел показаны на рисунке 1.
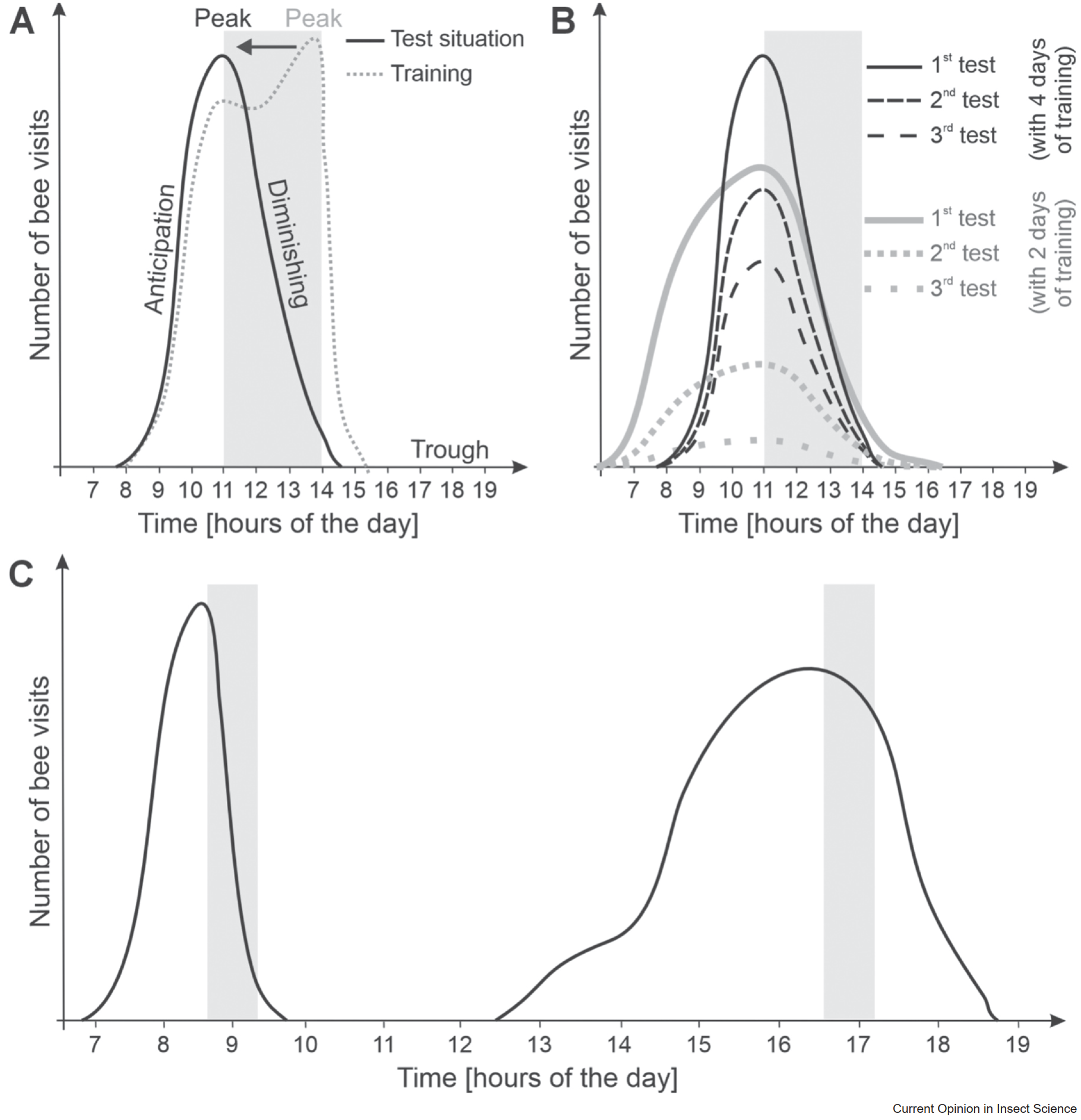
Рисунок 1. Как формируется пространственно-временная память у медоносных пчел. (A) В тестовых условиях, когда кормушка пуста (черная линия), активность пчел возрастает перед ожидаемым временем кормления (фаза предвкушения), достигая максимума в начале тренировочного периода (серые зоны), а затем резко снижается, когда пищи нет. Во время тренировок (серая пунктирная линия) посещаемость кормушки постепенно растет на протяжении всего периода кормления. (B) Опытные пчелы (черные кривые) точнее предугадывают время и дольше остаются активными у пустой кормушки в тестовые дни по сравнению с менее опытными (серые кривые). (C) Активность пчел зависит от времени суток: утренние посещения кормушки точно совпадают с тренировочным периодом, тогда как дневные посещения кормушки сильнее разбросаны по времени (ось X: с 6:00 до 19:00).
То, что пчелы прилетают к кормушке чуть раньше ожидаемого времени, не следует считать ошибкой их временной памяти. Напротив, исследования показывают, что пчелы продолжают посещать пустую кормушку в начале тренировочного периода в течение нескольких дней, причем их настойчивость зависит от опыта (Roy et al., 2025). Чем больше тренировочных дней или посещений у пчел, тем дольше сохраняется их память о времени кормления (Moore & Rankin, 1983; Moore et al., 2011). Кроме того, с увеличением числа тренировок пчелы становятся точнее в определении времени (Moore et al., 2011). Это может быть адаптацией к нестабильной доступности пищи в природе (Young & Dyer, 2022). Пчелы способны запомнить время кормления уже после одного дня тренировок (Beling, 1929; Moore et al., 2011), но при меньшем опыте они чаще совершают «проверочные» посещения кормушки вне тренировочного периода (Roy et al., 2025). Также показано, что фуражировочная активность пчел сильнее разбросана во второй половине дня (Moore & Rankin, 1983; Moore et al., 1989). Это может быть связано с менее предсказуемыми цветочными ресурсами или повышенной конкуренцией среди опылителей днем, что подтверждается увеличением числа опылителей на цветках масличных культур во второй половине дня (Karbassioon & Stanley, 2023) (Рисунок 1).
Также интересно узнать, связана ли способность пчел фуражировать в определенное время с внутренним механизмом отсчета времени. Белинг (Beling, 1929) и Валь (Wahl, 1932) показали, что даже при исключении всех известных внешних факторов пчелы сохраняют обученный ритм фуражирования (Beling, 1929; Wahl, 1932). Доказательство роли внутреннего механизма времени получил Реннер (Renner, 1955, 1957) в своем трансатлантическом эксперименте. Он перевез пчелиную колонию, обученную определенному времени кормления, из Парижа в Нью-Йорк за один день. Пчелы продолжали посещать кормушку в то же время, что и в Париже, несмотря на смену часового пояса, подтверждая наличие эндогенного механизма отсчета времени (Renner, 1955, 1957; Roy et al., 2025).
Интересно, что фуражировочные ритмы необученных пчел, не имеющих доступа к пище вне улья, похожи на их активность в лабораторных условиях (Kefuss & Nye, 1970; Moore & Rankin, 1985; Moore, 2001; Moore & Rankin, 1993). Эти ритмы, как и общая активность пчел, могут синхронизироваться с циклами света и темноты или режимами кормления (Frisch & Aschoff, 1987; Beier, 1968; Moore & Rankin, 1993). Возникает вопрос: связаны ли механизмы временной памяти и двигательной активности? Были предложены две гипотезы, объясняющие формирование временной памяти у пчел: 1) через систему осцилляторов, которая регулирует пороги возбуждения нейронов в центрах памяти; 2) через запоминание фазы центрального циркадного пейсмекера (CCP), связанной с определенным событием, так называемая «временная метка». Оба механизма зависят от CCP, который управляет суточной активностью пчел (Moore, 2001; Gallistel, 1990; Mistlberger, 1994). Первая гипотеза основана на идеях хронобиологии, а вторая — на исследованиях памяти и обучения (Roy et al., 2025).
Центральный циркадный пейсмекер (CCP), или главный часовой механизм в мозге пчел, получает информацию о времени суток через внешние сигналы, называемые Zeitgebers, в основном связанные со светом и температурой. Однако механические или химические сигналы также могут способствовать синхронизации этих часов (Helfrich-Forster, 2020; George & Stanewsky, 2021; Simoni et al., 2014; Ben-Shlomo & Kyriacou, 2002). Внутри темного, терморегулируемого улья химические сигналы играют важную роль в социальной синхронизации между пчелами (Siehler et al., 2021; Bloch, 2010) и, возможно, помогают формировать временную память через ограниченные циклы кормления. В клетках CCP происходят молекулярные колебания экспрессии генов внутренних часов, которые регулируют циркадные ритмы (Roy et al., 2025). Однако модель с одним осциллятором не объясняет способности пчел запоминать несколько временных периодов фуражирования в течение дня, что указывает на участие в формировании памяти дополнительных осцилляторов. В тканях пчел были обнаружены периферические (подчиненные) осцилляторы, которые работают автономно, но координируются CCP, влияя на поведение и физиологические ритмы (Bell-Pedersen et al., 2005; Roy et al., 2025).
Модель с множественными осцилляторами, включающая один центральный осциллятор, реагирующий на свет, и периферические, связанные с циклами кормления, была предложена для пространственно-временного обучения (TPL) у млекопитающих (крыс, хомяков и мышей) (Mistlberger, 1994; Mulder et al., 2013; Ralph et al., 2013). Однако для пчел такая модель не подходит, так как они способны запоминать до девяти временных периодов кормления в день (Koltermann, 1971). Это потребовало бы координации до девяти периферических осцилляторов с CCP, что представляется маловероятным (Moore, 2001; Mistlberger, 1994). Ежедневные изменения в предвосхищающем поведении пчел также указывают на отсутствие множества равнозначных периферических осцилляторов (Moore & Rankin, 1983; Moore et al., 2001; Roy et al., 2025). Кроме того, способность пчел фуражировать у двух кормушек в разное время (Wahl, 1932) или связывать разные цвета с определенным временем суток (Zhang et al., 2006) сложно объяснить только через вовлечение осцилляторов, что поддерживает гипотезу о «временной метке» — запоминании фазы циркадного цикла, связанной с событием.
Исследования Паля и др. (Pahl et al., 2007) показали, что пчелы могут формировать сложные ассоциации между визуальными узорами, местоположением и временем суток, объединяя компоненты «что», «где» и «когда». Это напоминает эпизодическую память, характерную для позвоночных (Babb & Crystal, 2006; Clayton & Dickinson, 1998; Lewis et al., 2003). У пчел такое поведение регулируется циркадным временем, а не способностью измерять интервалы между событиями, как у позвоночных, поэтому его называют «циркадно-временной эпизодической памятью» (Koltermann, 1971). Однако шмели (Bombus impatiens), близкие родственники пчел, способны измерять интервалы между событиями (Boisvert & Sherry, 2006), и это применимо к медоносным пчелам (Roy et al., 2025). Чтобы выяснить, как сильно циркадные часы контролируют фуражировочную активность пчел, нужны эксперименты с манипуляцией генов часов или повреждением CCP.
Первые данные о молекулярных основах временной памяти были получены на модельных животных с встроенными генетическими инструментами. Например, у линий мышей с отключенными генами криптохрома (m-cry, аналогом которого у насекомых является криптохром 2) нарушалась способность запоминать и воспроизводить информацию о времени суток (Van der Zee et al., 2008). Аналогично, у мушек дрозофил с мутациями в генах period (per) и clock (Clk) наблюдались проблемы с обучением, связанным с временем (Chouhan et al., 2015). Интересно, что у мышей гены per1 и per2 не обязательны для пространственно-временного обучения (TPL), но необходимы для нормальной двигательной активности. Это позволило предположить, что временная память у млекопитающих зависит не от генов per, а от гена m-cry, связанного с периферическим осциллятором (Mulder et al., 2013).
Изучение молекулярных механизмов временной памяти у медоносных пчел началось сравнительно недавно (Naeger et al., 2011). В одном из первых систематических экспериментов пчел из одной колонии обучали посещать две кормушки — одну утром, другую днем. Анализ мозга с помощью микрочипов выявил 1329 генов, экспрессия которых различалась между пчелами, ожидающими пищу, и неактивными пчелами. Это указывало на связь временной памяти с генетическими изменениями в мозге. Например, у пчел в состоянии предвкушения повышалась экспрессия генов, связанных с анкирином и рецепторами инсулина, тогда как гены часов per и cry2 были более активны у неактивных пчел, что соответствует их суточным ритмам (Rubin et al., 2006). Позже было установлено, что ген Egr1 (ранний белок роста 1) это транскрипционный фактор, связанный с обучением и регулируемый нейронной активностью (Lutz & Robinson, 2013; Ugajin et al., 2013), играет важную роль в формировании временной памяти пчел. Было показано, что экспрессия гена Egr1 и связанных с ним генов, таких как рецептор гормона 38, увеличивается в мозге фуражирующих пчел во время обучения на кормушке (Singh et al., 2018). Изменения в экспрессии генов Egr1, per и cry2 после активации процессов временной памяти показаны на рисунке 2.
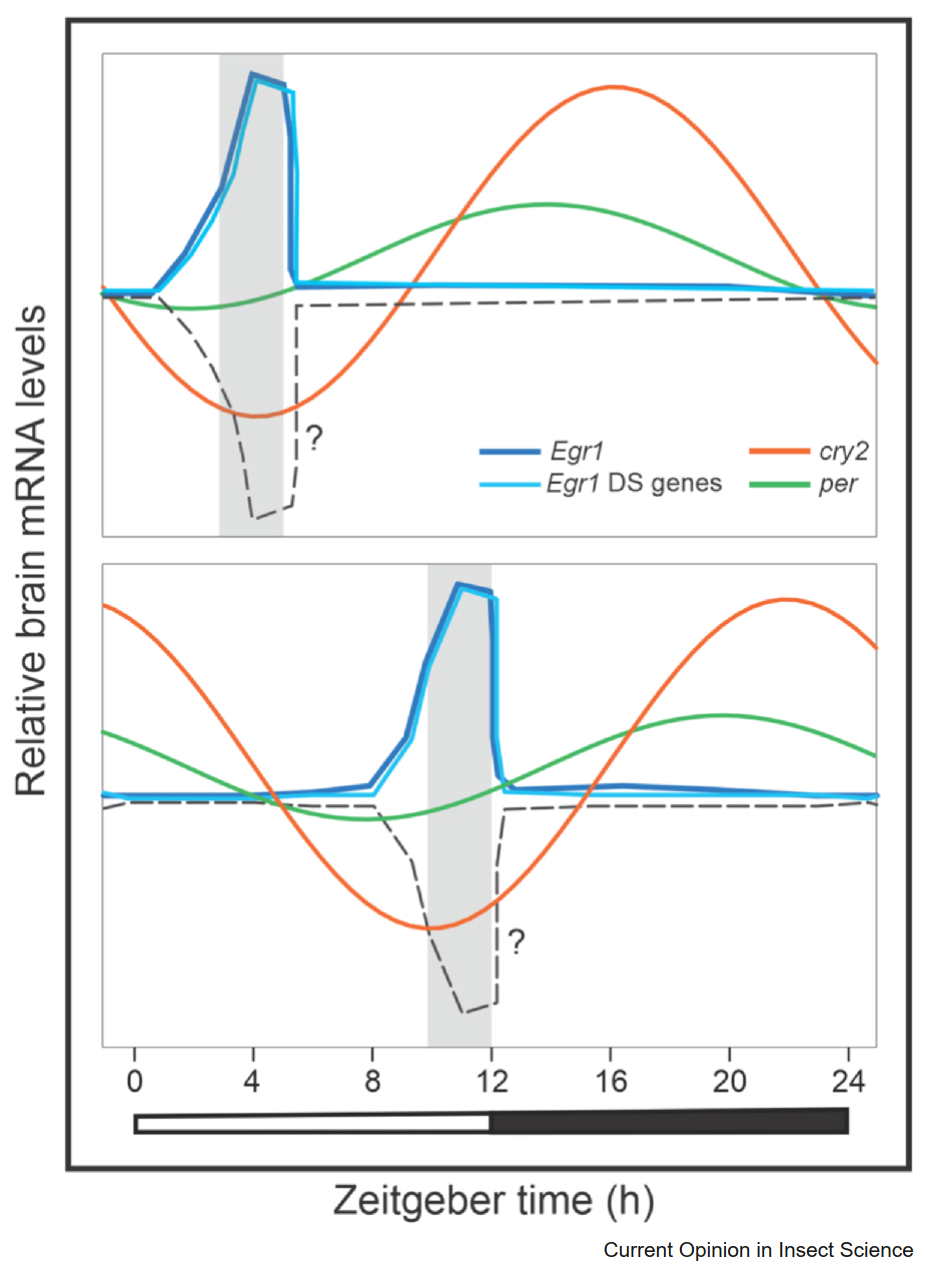
Рисунок 2. Изменения экспрессии генов Egr1, per и cry2 при формировании временной памяти у пчел. Анализ генов в мозге пчел показал, что экспрессия гена Egr1 (темно-синяя линия) и связанных с ним генов (светло-синяя линия), таких как Hr38 и рецептор дофамина и экдистероидов (DopEcR), достигает пика во время обучения. Экспрессия основных часовых генов cry2 (оранжевая линия) и per (зеленая линия) также связана с временем обучения (серые затененные области). Пунктирная линия отражает гены, чья экспрессия строго зависит от времени обучения, но минимальна в момент обучения. К таким генам могут относиться те, что участвуют в метаболизме, мембранном транспорте или подавлении транскрипции. Верхняя и нижняя панели показывают два разных времени обучения: в обоих случаях экспрессия Egr1 и его связанных генов резко возрастает во время обучения, тогда как экспрессия часовых генов cry2 и per меняется в зависимости от режима обучения.
Повышение экспрессии гена Egr1 у обученных фуражирующих пчел наблюдалось даже при искусственном предотвращении их вылета, например, с помощью имитации дождя (Shah et al., 2018). Это говорит о том, что экспрессия Egr1 может регулироваться внутренними сигналами циркадных часов. У шмелей также отмечено увеличение экспрессии генов Egr1 и Hr38 во время фуражировочных полетов (Iino et al., 2020). Однако для полного понимания молекулярных механизмов временной памяти необходим более детальный анализ экспрессии генов в нейронных популяциях, связанных с этими процессами. Одним из основных регионов памяти мозга являются малые клетки Кеньона в грибовидных телах, которые показывают повышенную нейронную активность при фуражировке (Lutz & Robinson, 2013; Iino et al., 2020) и временном обучении (Shah et al., 2018). Кроме того, временное обучение влияет на молекулярные часы в трех тканях пчел — во всем мозге, подглоточном ганглии и антеннах (Jain & Brockmann, 2018), что показывает связь между циркадными часами и временной памятью. Эти исследования указывают, что временная память сопровождается изменениями в экспрессии часовых генов в различных нейронных тканях и других генов на уровне всего мозга, вероятно, под контролем молекулярных часов (Рисунок 2).
Чтобы информация о времени суток передавалась в центры памяти, позволяя пчелам вспоминать контекстные детали в нужный момент, необходимы функциональные связи. Существует два наиболее вероятных механизма: 1) прямые сигналы от центрального циркадного пейсмекера (CCP) через рецепторы циркадных нейропептидов; 2) наличие локальных часовых клеток в центрах памяти. Около 50 лет назад Мартин и др. (Martin et al., 1978) провели эксперимент по трансплантации грибовидных тел у пчел, обученных разным временным режимам. После пересадки профили фуражировочной активности реципиентов соответствовали режиму донора, что позволило предположить гуморальный перенос информации о времени (Martin et al., 1978). Аналогично, исследования на тараканах показали, что удаление CCP не влияло на обонятельное обучение, но нарушило способность формировать и извлекать память в зависимости от времени суток в экспериментах с классическим и оперантным обусловливанием (Lubinski & Page, 2016). У крыс повреждение CCP также блокировало зависимое от времени формирование памяти (Stephan & Kovacevic, 1978). У медоносных пчел эффективность обонятельного обучения зависит от фазы суточного цикла, и это не просто реакция на темноту ночью, так как циркадный паттерн памяти сохраняется в постоянных условиях (Lehmann et al., 2011). Кроме того, извлечение цветовой памяти у пчел также зависит от циркадной фазы (Prabhu & Cheng, 2008; Gerstner & Yin, 2010; Roy et al., 2025).
Однако точные связи между циркадными часами и центрами памяти у пчел еще не установлены. Часовые нейроны, экспрессирующие гены Period (PER) и Pigment Dispersing Factor (PDF) — нейропептид, производимый только циркадными нейронами, — расположены близко к грибовидным телам (Beer et al., 2018; Fuchikawa et al., 2017). Это создает возможность передачи информации о времени суток через паракринную сигнализацию PDF. Исследования на дрозофилах показали, что PDF-содержащие нейроны демонстрируют циркадную пластичность, зависящую от сигнализации PDF (Fernandez et al., 2008; Herrero et al., 2020). Схема связей между CCP и центрами памяти в мозге медоносных пчел представлена на схеме (Рисунок 3).
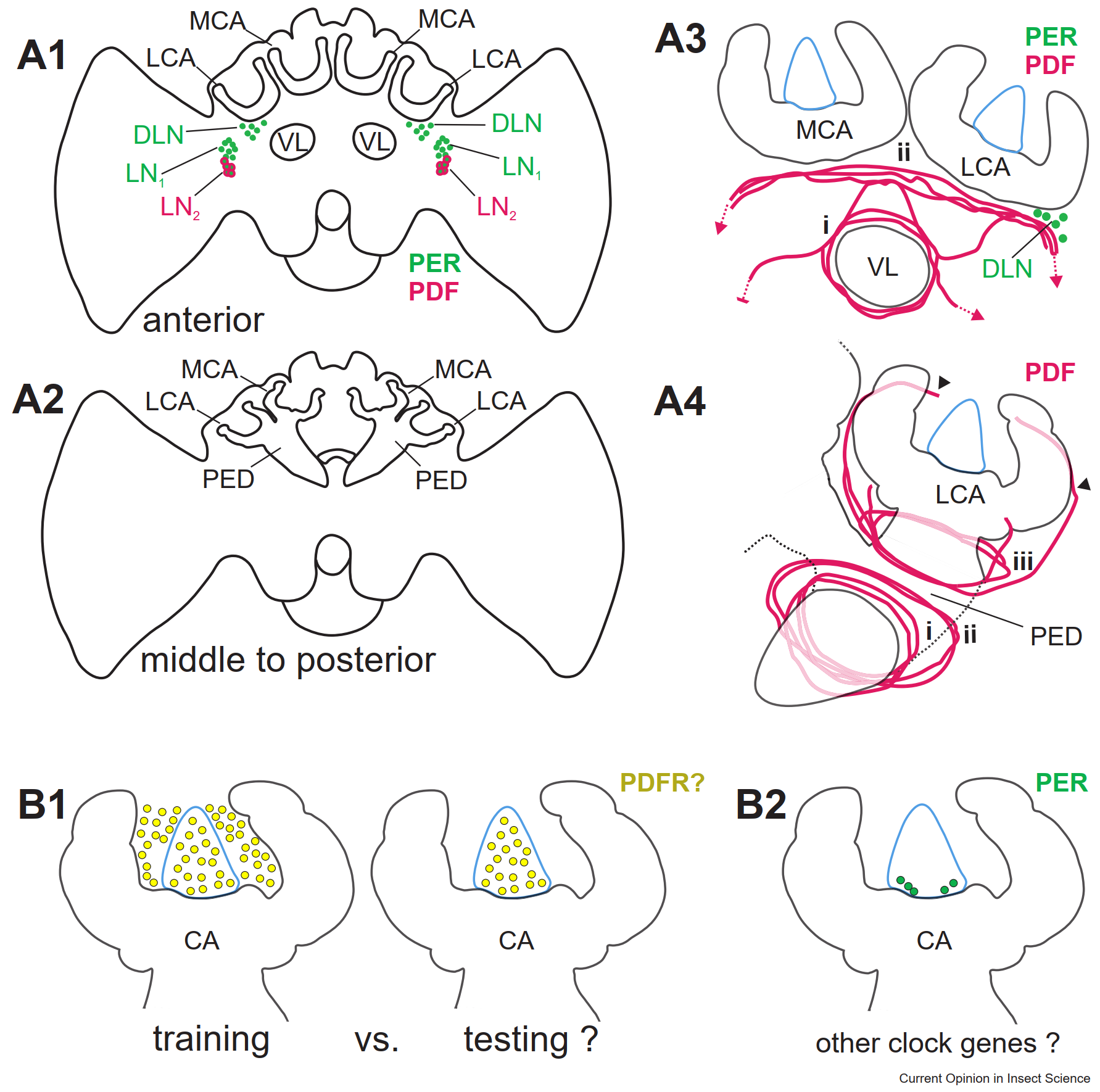
Рисунок 3. Схема связей между центральным циркадным пейсмекером (CCP) и центрами памяти в мозге медоносных пчел. (A) Нейроны, экспрессирующие ген нейропептида PDF (кластер LN2, показан пурпурным), находятся в CCP (схемы передней [A1] и средней/задней [A2] частей мозга). Эти нейроны также экспрессируют ген часов PER (зеленый) и связаны с другими кластерами нейронов (LN1, DLN) [A3]. Вместе они образуют сеть, охватывающую грибовидные тела (MBs) [A3, A4]. В передней части мозга волокна PDF (i) окружают вертикальную долю грибовидных тел (VL), а другой пучок (ii) проходит рядом с чашечками (MCA, LCA) [A3]. В средней и задней частях грибовидных тел (PED) окружен сетью волокон (i, ii), формирующей кольцо у чашечек (iii) с разветвлениями назад (черные стрелки). Сеть PDF, показанная для PED и LCA [A4], аналогична для MCA. (B) Изменения экспрессии гена рецептора PDF (PDFR) в грибовидных телах во время тренировок и тестов на временную память (аналогично гену Egr1) указывают на сигналы, поступающие от CCP [B1]. Экспрессия гена PER у основания чашечек грибовидных тел может свидетельствовать о наличии локального осциллятора [B2], если будет подтверждена совместная экспрессия других часовых генов и их циркадные колебания. CA — чашечка грибовидных тел; субпопуляция клеток Кеньона обозначена синим.
Аксоны часовых нейронов в спинном мозге дрозофил изменяются в течение суток, образуя больше терминалей днем, чем ночью (Fernandez et al., 2008). Также была обнаружена косвенная связь между центральным циркадным пейсмекером (CCP) и грибовидными телами (MBs) через рецепторы PDF (PDFR) в интернейронах, расположенных за пределами MBs (Flyer-Adams et al., 2020). Недавний анализ коннектома дрозофил подтвердил, что связи между CCP и MBs преимущественно косвенные (Reinhard et al., 2023). Эти данные подтвердили, что CCP передает сигналы в центры памяти насекомых. Если бы рецепторы PDFR были найдены в клетках Кеньона — нейронах грибовидных тел, — это стало бы доказательством прямой сигнализации от CCP (Рисунок 3) (Roy et al., 2025). У позвоночных клетки гиппокампа, центра памяти, экспрессируют гены часов, что указывает на их роль как локальных осцилляторов (Shieh, 2003; Reinhard et al., 2023). У насекомых такие клетки пока не идентифицированы, но Фучикава и др. (Fuchikawa et al., 2017) нашли предварительные признаки локальных часов в центрах памяти пчел: клетки, экспрессирующие ген PER, расположены у основания чашечек грибовидных тел. Однако для подтверждения их роли как функционального осциллятора требуется доказать совместную экспрессию других часовых генов и их циркадные колебания (Fuchikawa et al., 2017).
Временная память пчел напоминает эпизодическую память позвоночных, а их молекулярные часы схожи с часами млекопитающих: у перепончатокрылых есть ген криптохрома типа млекопитающих (cry2), но отсутствует ген насекомых cry1 (Pahl et al., 2007; Rubin et al., 2006). Это предполагает на сходство в механизмах регуляции. Однако есть и различия: эпизодическая память пчел привязана к циркадному времени (Pahl et al., 2007), и они способны запоминать до девяти временных периодов в день (Koltermann, 1971), тогда как позвоночные обычно запоминают только два-четыре периода (Biebach et al., 1989; Van der Zee et al., 2008; Reebs, 1996; Clayton & Dickinson, 1998). Преимущество пчел в изучении механизмов формирования временной памяти в том, что их сложные процессы памяти регулируются относительно простым набором часовых генов — у насекомых только один ген per против трех у млекопитающих (Koltermann, 1971; Mulder et al., 2013). Кроме того, анатомические исследования выявляют разнообразные связи между внутренними часами и центрами памяти у пчел, которые сложно изучать у дрозофил, так как у них отсутствует сложное пространственно-временное поведение TPL (Beer & Helfrich-Forster, 2020). Перепончатокрылые, включая пчел, представляют собой группу с разными образами жизни (социальными и одиночными), диетами и адаптациями, что делает их ценными моделями для исследования эволюции формирования временной памяти (Roy et al., 2025).
Список литературы
- Babb SJ, Crystal JD: Discrimination of what, when, and where is not based on time of day. Learn Behav 2006, 34:124-130, https:// doi.org/10.3758/BF03193188
- Beer K, Helfrich-Forster C: Model and non-model insects in chronobiology. Front Behav Neurosci 2020, 14:1-23, https://doi. org/10.3389/fnbeh.2020.601676
- Beer K, Kolbe E, Kahana NB, Yayon N, Weiss R, Menegazzi P, Bloch G, •• Helfrich-Forster C: Pigment-dispersing factor-expressing neurons convey circadian infoBeer K, Zupanc GKH, Helfrich-Forster C: Ingeborg Beling and the time memory in honeybees: almost one hundred years of research. J Comp Physiol A 2024, 210:189-201, https://doi.org/10. 1007/s00359-024-01691-9rmation in the honey bee brain. Open Biol 2018, 8:1-24, https://doi.org/10.1098/rsob.170224.
- Beier W, Lindauer M: Der Sonnenstand als Zeitgeber fur die Biene. Apidologie 1970, 1:5-28, https://doi.org/10.1051/ apido:19700101
- Beier W: Beeinflussung der inneren Uhr der Bienen durch Phasenverschiebung des Licht-Dunkel-Zeitgebers. Z Fur Bienenforsch 1968, 9:356-378.
- Beling I: Uber das Zeitgedachtnis der bienen (https://doi.org/ https:://doi.org/), Z Fur Vgl Physiol 1929, 9:259-338, https://doi.org/ 10.1007/BF00340159
- Bell-Pedersen D, Cassone VM, Earnest DJ, Golden SS, Hardin PE, Thomas TL, Zoran MJ: Circadian rhythms from multiple oscillators: lessons from diverse organisms. Nat Rev Drug Discov (7) 2005, 6:544-556, https://doi.org/10.1038/nrd1633
- Ben-Shlomo R, Kyriacou CP: Circadian rhythm entrainment in flies and mammals. Cell Biochem Biophys 2002, 37:141-156, https://doi.org/10.1385/CBB:37:2:141
- Biebach H, Gordijn M, Krebs JR: Time-and-place learning by garden warblers, Sylvia borin. Anim Behav 1989, 37:353-360, https://doi.org/10.1016/0003-3472(89)90083-3
- Bloch G, Bar-Shai N, Cytter Y, Green R: Time is honey: circadian clocks of bees and flowers and how their interactions may influence ecological communities. Philos Trans R Soc B Biol Sci 2017, 372:20160256, https://doi.org/10.1098/rstb.2016.0256
- Bloch G: The social clock of the honeybee. J Biol Rhythms 2010, 25:307-317, https://doi.org/10.1177/0748730410380149
- Boisvert MJ, Sherry DF: Interval timing by an invertebrate, the bumble bee Bombus impatiens. Curr Biol 2006, 16:1636-1640, https://doi.org/10.1016Zj.cub.2006.06.064
- Chouhan NS, Wolf R, Helfrich-Forster C, Heisenberg M: Flies remember the time of day. Curr Biol 2015, 25:1619-1624, https:// doi.org/10.1016/j.cub.2015.04.032
- Clayton NS, Dickinson A: Episodic-like memory during cache recovery by scrub jays. Nature 1998, 395:272-274, https://doi. org/10.1038/26216
- Elizalde L, Arbetman M, Arnan X, Eggleton P, Leal IR, Lescano MN, Saez A, Werenkraut V, Pirk GI: The ecosystem services provided by social insects: traits, management tools and knowledge gaps. Biol Rev 2020, 95:1418-1441, https://doi.org/10.1111/brv.12616
- Fernandez MP, Berni J, Ceriani MF: Circadian remodeling of neuronal circuits involved in rhythmic behavior. PLoS Biol 2008, 6:e69.
- Flyer-Adams JG, Rivera-Rodriguez EJ, Yu J, Mardovin JD, Reed •• ML, Griffith LC: Regulation of olfactory associative memory by the circadian clock output signal Pigment-Dispersing Factor (PDF). J Neurosci 2020, 40:9066-9077, https://doi.org/10.1523/ JNEUR0SCI.0782-20.2020.
- Frisch B, Aschoff J: Circadian rhythms in honeybees: entrainment by feeding cycles. Physiol Entomol 1987, 12:41-49, https://doi.org/10.1111/j.1365-3032.1987.tb00722.x
- Fuchikawa T, Beer K, Linke-Winnebeck C, Ben-David R, Kotowoy •• A, Tsang VWK, Warman GR, Winnebeck EC, Helfrich-Forster C, Bloch G: Neuronal circadian clock protein oscillations are similar in behaviourally rhythmic forager honeybees and in arrhythmic nurses. Open Biol 2017, 7:1-20, https://doi.org/10. 1098/rsob.170047.
- Gallistel CR: The Organization of Learning. The MIT Press; 1990.
- George R, Stanewsky R: Peripheral sensory organs contribute to temperature synchronization of the circadian clock in Drosophila melanogaster. Front Physiol 2021, 12:622545, https:// doi.org/10.3389/fphys.2021.622545
- Gerstner JR, Yin JCP: Circadian rhythms and memory formation. Nat Rev Neurosci 2010, 11:577-588, https://doi.org/10.1038/nrn2881
- Gimenes M, Benedito-Silva AA, Marques MD: Circadian rhythms of pollen and nectar collection by bees on the flowers of Ludwigia elegans (Onagraceae). Biol Rhythm Res 1996, 27:281-290, https://doi.org/10.1076/brhm.27.3.281.12971
- Gonulkirmaz-Cancalar O, Shertzer O, Bloch G: Bumble Bees (Bombus terrestris) use time-memory to associate reward with color and time of day. Insects 2023, 14:707, https://doi.org/10. 3390/insects14080707.
- Harrison JM, Breed MD: Temporal learning in the giant tropical ant, Paraponera clavata. Physiol Entomol 1987, 12:317-320, https://doi.org/10.1111/j.1365-3032.1987.tb00756.x
- Helfrich-Forster C: Light input pathways to the circadian clock of insects with an emphasis on the fruit fly Drosophila melanogaster. J Comp Physiol A 2020, 206:259-272, https://doi. org/10.1007/s00359-019-01379-5
- Herrero A, Yoshii T, Ispizua Jl, Colque C, Veenstra JA, Muraro Nl, • Ceriani MF: Coupling neuropeptide levels to structural plasticity in Drosophila clock neurons, e4, Curr Biol 2020, 30:3154-3166, https://doi.org/10.1016/j.cub.2020.06.009.
- Jain R, Brockmann A: Time-restricted foraging under natural •• light/dark condition shifts the molecular clock in the honey bee, Apis mellifera. Chronobiol Int 2018, 35:1723-1734, https:// doi.org/10.1080/07420528.2018.1509867.
- Karbassioon A, Stanley DA: Exploring relationships between time of day and pollinator activity in the context of pesticide use. Basic Appl Ecol 2023, 72:74-81, https://doi.org/10.1016/j. baae.2023.06.001
- Kefuss JA, Nye WP: The influence of photoperiod on the flight activity of honeybees. J Apic Res 1970, 9:133-139, https://doi. org/10.1080/00218839.1970.11100260
- Kleber E: Hat das Zeitgedachtnis der Bienen Biologische Bedeutung? Z Fur Vgl Physiol 1935, 22:221-262, https://doi.org/ 10.1007/BF00586500
- Koltermann R: 24-Std-Periodik in der Langzeiterinnerung an Duft- und Farbsignale bei der Honigbiene. Z Fur Vgl Physiol 1971, 75:49-68, https://doi.org/10.1007/BF00335137
- Lehmann M, Gustav D, Galizia CG: The early bee catches the flower — circadian rhythmicity influences learning performance in honey bees, Apis mellifera. Behav Ecol Sociobiol 2011, 65:205-215, https://doi.org/10.1007/s00265-010-1026-9
- Lewis PA, Miall RC, Daan S, Kacelnik A: Interval timing in mice does not rely upon the circadian pacemaker. Neurosci Lett 2003, 348:131-134, https://doi.org/10.1016/S0304-3940(03)00521-4
- Lubinski AJ, Page TL: The optic lobes regulate circadian rhythms of olfactory learning and memory in the cockroach. J Biol Rhythms 2016, 31:161-169, https://doi.org/10.1177/ 0748730415622710
- Lutz CC, Robinson GE: Activity-dependent gene expression in honey bee mushroom bodies in response to orientation flight. J Exp Biol 2013, 216:2031-2038, https://doi.org/10.1242/jeb.084905
- Martin U, Martin H, Lindauer M: Transplantation of a time-signal in honeybees. J Comp Physiol A 1978, 124:193-201, https://doi. org/10.1007/BF00657051
- Medugorac I, Lindauer M: Das Zeitgedachtnis der Bienen unter dem EinfluB von Narkose und von sozialen Zeitgebern. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 1967, 55:450-474, https://doi.org/10.1007/BF00302625
- Mistlberger RE: Circadian food-anticipatory activity: formal models and physiological mechanisms. Neurosci Biobehav Rev 1994, 18:171-195, https://doi.org/10.1016/0149-7634(94)90023-X
- Moore D, Doherty P: Acquisition of a time-memory in forager honey bees. J Comp Physiol A 2009, 195:741-751, https://doi.org/ 10.1007/s00359-009-0450-7
- Moore D, Rankin MA: Circadian locomotor rhythms in individual honeybees. Physiol Entomol 1985, 10:191-197, https://doi.org/10. 1111/j.1365-3032.1985.tb00034.x
- Moore D, Rankin MA: Diurnal changes in the accuracy of the honeybee foraging rhythm. Biol Bull 1983, 164:471-482, https:// doi.org/10.2307/1541256
- Moore D, Rankin MA: Light and temperature entrainment of a locomotor rhythm in honeybees. Physiol Entomol 1993, 18:271-278, https://doi.org/10.1111/j.1365-3032.1993.tb00599.x
- Moore D, Siegfried D, Wilson R, Rankin MA: The influence of time of day on the foraging behavior of the honeybee, Apis mellifera. J Biol Rhythms 1989, 4:305-325, https://doi.org/10.1177/ 074873048900400301
- Moore D, Van Nest BN, Seier E: Diminishing returns: the influence of experience and environment on time-memory extinction in honey bee foragers. J Comp Physiol A 2011, 197:641-651, https://doi.org/10.1007/s00359-011-0624-y
- Moore D: Honey bee circadian clocks: behavioral control from individual workers to whole-colony rhythms. J Insect Physiol 2001, 47:843-857, https://doi.org/10.1016/S0022-1910(01)00057-9
- Mulder C, Van Der Zee EA, Hut RA, Gerkema MP: Time-place • learning and memory persist in mice lacking functional peri and per2 clock genes. J Biol Rhythms 2013, 28:367-379, https:// doi.org/10.1177/0748730413512958.
- Murphy CM, Breed MD: Time-place learning in a neotropical stingless bee, Trigona fulviventris guerin (Hymenoptera: Apidae). J Kans Entomol Soc 2008, 81:73-76, https://doi.org/10. 2317/JKES-704.23.1
- Naeger NL, Van Nest BN, Johnson JN, Boyd SD, Southey BR, Rodriguez-Zas SL, Moore D, Robinson GE: Neurogenomic signatures of spatiotemporal memories in time-trained forager honey bees. J Exp Biol 2011, 214:979-987, https://doi.org/10. 1242/jeb.053421.
- Ofstad TA, Zuker CS, Reiser MB: Visual place learning in Drosophila melanogaster. Nature 2011, 474:204-207, https://doi. org/10.1038/nature10131
- Pahl M, Zhu H, Pix W, Tautz J, Zhang S: Circadian timed episodic-like memory a bee knows what to do when, and also where. J Exp Biol 2007, 210:3559-3567, https://doi.org/10.1242/jeb.005488.
- Prabhu C, Cheng K: One day is all it takes: circadian modulation of the retrieval of colour memories in honeybees. Behav Ecol Sociobiol 2008, 63:11-22, https://doi.org/10.1007/s00265-008-0631-3
- Ralph MR, Sam K, Rawashdeh OA, Cain SW, Ko CH: Memory for time of day (time memory) is encoded by a circadian oscillator and is distinct from other context memories. Chronobiol Int 2013, 30:540-547, https://doi.org/10.3109/07420528.2012.754449
- Reebs SG: Time-place learning in golden shiners (Pisces: Cyprinidae). Behav Process 1996, 36:253-262, https://doi.org/10. 1016/0376-6357(96)88023-5
- Reinhard J, Srinivasan MV, Guez D, Zhang SW: Floral scents induce recall of navigational and visual memories in honeybees. J Exp Biol 2004, 207:4371-4381, https://doi.org/10. 1242/jeb.01306
- Reinhard J, Srinivasan MV, Zhang S: Complex memories in honeybees: can there be more than two? J Comp Physiol A 2006, 192:409-416, https://doi.org/10.1007/s00359-005-0079-0
- Reinhard J, Srinivasan MV, Zhang S: Scent-triggered navigation in honeybees. Nature 2004, 427:411-411, https://doi.org/10.1038/ 427411a
- Reinhard, N; Fukuda, A; Manoli, G; Derksen, E; Saito, A; Moller, G; Sekiguchi, M; Rieger, D; Helfrich-Forster, C; Yoshii, T; Zandawala M : Synaptic Connectome of the Drosophila Circadian Clock; 2023. (https://doi.org/10.1101/2023.09.11.55722).
- Renner M: Ein Transozeanversuch zum Zeitsinn der Honigbiene. Naturwissenschaften 1955, 42:540-541, https://doi. org/10.1007/BF00630156
- Renner M: Neue Versuche uber den Zeitsinn der Honigbiene. Z Fur Vgl Physiol 1957, 40:85-118, https://doi.org/10.1007/BF00298152
- Renner M: Uber Ein weiteres Versetzungsexperiment zur Analyse des Zeitsinnes und der Sonnenorientierung der Honigbiene. Z Fur Vgl Physiol 1959, 42:449-483, https://doi.org/ 10.1007/BF00297804
- Roy, T., Beer, K., 2025. Time memory in social insects with a special focus on honey bees. Curr Opin Insect Sci. 68, 101327. doi: 10.1016/j.cois.2024.101327.
- Rubin EB, Shemesh Y, Cohen M, Elgavish S, Robertson HM, Bloch G: Molecular and phylogenetic analyses reveal mammalian-like clockwork in the honey bee (Apis mellifera) and shed new light on the molecular evolution of the circadian clock. Genome Res 2006, 16:1352-1365, https://doi.org/10.1101/gr.5094806
- Schatz B, Beugnon G, Lachaud J-P: Time-place learning by an invertebrate, the ant Ectatomma ruidum Roger. Anim Behav 1994, 48:136-238, https://doi.org/10.1006/anbe.1994.1232
- Shah A, Jain R, Brockmann A: Egr-1: a candidate transcription •• factor involved in molecular processes underlying time- memory. Front Psychol 2018, 9:865, https://doi.org/10.3389/fpsyg. 2018.00865.
- Shieh K-R: Distribution of the rhythm-related genes rPERIOD1, rPERIOD2, and rCLOCK, in the rat brain. Neuroscience 2003, 118:831-843, https://doi.org/10.1016/S0306-4522(03)00004-6
- Siehler O, Wang S, Bloch G: Remarkable sensitivity of young honey bee workers to multiple non-photic, non-thermal, forager cues that synchronize their daily activity rhythms. Front Physiol 2021, 12:789773, https://doi.org/10.3389/fphys.2021.789773
- Simoni A, Wolfgang W, Topping MP, Kavlie RG, Stanewsky R, Albert JT: A Mechanosensory pathway to the Drosophila circadian clock. Science 2014, 343:525-528, https://doi.org/10. 1126/science.1248186
- Singh AS, Shah A, Brockmann A: Honey bee foraging induces •• upregulation of Early Growth Response Protein 1, Hormone Receptor 38 and candidate downstream genes of the ecdysteroid signalling pathway. Insect Mol Biol 2018, 27:90-98, https://doi.org/10.1111/imb.12350.
- Stephan FK, Kovacevic NS: Multiple retention deficit in passive avoidance in rats is eliminated by suprachiasmatic lesions. Behav Biol 1978, 22:456-462, https://doi.org/10.1016/S0091- 6773(78)92565-8
- Toure, MW; Young, FJ; McMillan, WO; Montgomery, SH : Heliconiini Butterflies Can Learn Time-Dependent Reward Associations; (http://dx.doi.org/10.1098/rsbl.2020.0424).
- Ugajin A, Kunieda T, Kubo T: Identification and characterization of an Egr ortholog as a neural immediate early gene in the European honeybee (Apis mellifera L.). FEBS Lett 2013, 587:3224-3230, https://doi.org/10.1016/j.febslet.2013.08.014
- Van der Zee EA, Havekes R, Barf RP, Hut RA, Nijholt IM, Jacobs EH, Gerkema MP: Circadian time-place learning in mice depends on cry genes. Curr Biol 2008, 18:844-848, https://doi. org/10.1016/j.cub.2008.04.077
- Wahl O: Neue Untersuchungen uber das Zeitgedachtnis der Bienen. Z Fur Vgl Physiol 1932, 16:529-589, https://doi.org/10. 1007/BF00338333
- Young AM, Dyer FC: Past experience with spatial or temporal resource unpredictability shapes exploration in honey bees, Apis mellifera. Anim Behav 2022, 194:253-264, https://doi.org/10. 1016/j.anbehav.2022.09.001
- Zhang S, Schwarz S, Pahl M, Zhu H, Tautz J: Honeybee memory: a honeybee knows what to do and when. J Exp Biol 2006, 209:4420-4428, https://doi.org/10.1242/jeb.02522
- de Jesus TNCS, Venturieri GC, Contrera FAL: Time-place learning in the bee Melipona fasciculata (Apidae, Meliponini). Apidologie 2014, 45:257-265, https://doi.org/10.1007/s13592-013-0245-2
- lino S, Shiota Y, Nishimura M, Asada S, Ono M, Kubo T: Neural activity mapping of bumble bee (Bombus ignitus) brains during foraging flight using Immediate early genes. Sci Rep 2020, 10:7887, https://doi.org/10.1038/s41598-020-64701-1
Новость подготовили
© д.б.н., в.н.с. Ильясов Рустем Абузарович, Ильясова Алла Юрьевна
01.11.2025