 © Белкина Елена Геннадьевна,
младший научный сотрудник лаборатории эволюционной генетики развития ИБР РАН.
© Белкина Елена Геннадьевна,
младший научный сотрудник лаборатории эволюционной генетики развития ИБР РАН.
Область научных интересов: Drosophila, брачное поведение, поведенческие программы, изолирующие барьеры, генетика поведения, эволюция.
Репродуктивное поведение D. melanogaster представляет собой обмен сигналами различной модальности между потенциальными брачными партнерами. Различают визуальные, тактильные, акустические и химические способы получения и передачи информации. Акустический канал является одним из наиболее изученных каналов коммуникации. Самцы Drosophila генерируют видоспецифичную брачную песню путем отведения крыла в сторону и вибрации им на определенной частоте. Инициация акустического поведения самца контролируется двумя генами транскрипционных факторов, определяющих пол у дрозофилы – fruitless (fru) и doublesex (dsx). Различия в их экспрессии определяют формирование как морфологических, так и поведенческих половых признаков. Оптогенетическая и термогенетическая активация как fru нейронов, так и dsx нейронов у самцов приводит к генерации брачной песни даже в отсутствие самки. Инициация акустического сигнала самца происходит по нижеследующей схеме. После восприятия сигналов различных модальностей, генерация песни запускается нейронами кластера мозга P1, откуда информация передается на нисходящие нейроны P2b и pIP10, которые в свою очередь посылают аксоны в торакальный ганглий. dPR1 (нейроны в проторакальном ганглии) и vPR6 (нейроны в мезоторакальном ганглии) представляют собой компоненты центрального генератора моторного паттерна песни, в функции которого входят отведение крыла и звукогенерация. За продуцирование песни ответственны два вида крыловых мышц, а именно, мышцы прямого и непрямого действия. Крыловые мышцы прямого действия отвечают за структуру акустического сигнала, тогда как мышцы непрямого действия – за амплитуду исполняемой брачной песни.
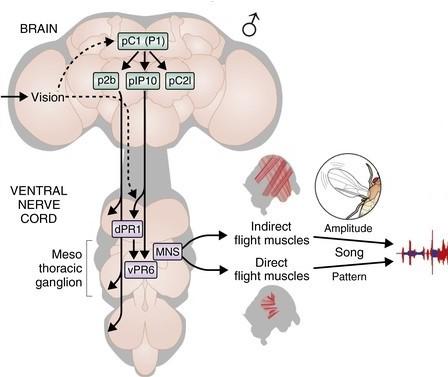
Рис. 1. Инициация акустического поведения самца в мозгу и брюшной нервной цепи. Рисунок из Coen & Murthy, 2016. https://doi.org/10.1016/j.conb.2016.01.013
Долгое время считалось, что в брачной паре D. melanogaster только самец исполняет песню, тогда как самка является лишь безмолвным «слушателем», объектом ухаживаний самца; но за которой, тем не менее, остается право окончательного выбора. Новое исследование, выполненное учеными из университета Орхуса (Дания), показало способность самок генерировать во время спаривания акустический сигнал, отличный от брачной песни самца (исполняемой во время ухаживания, до копуляции). Без сомнения, эта скрупулезно выполненная работа предлагает открытие новых горизонтов в изучении акустического дуэта в контексте брачного поведения.
Копуляционная песня самки и брачная песня самца имеют ряд отличий. Во-первых, песня самки предполагает одновременное использование обоих крыльев, тогда как песня самца – использование одного крыла. Во-вторых, есть отличия в песенных параметрах. В частности, песня самки состоит из множества повторяющихся циклов высокой частоты и более длительными интервалами между ними. В-третьих, в отличие от видоспецифичной брачной песни самца, песня самки является полоспецифичной, что показано авторами на близкородственных видах D. melanogaster, D. simulans и D. mauritiana.
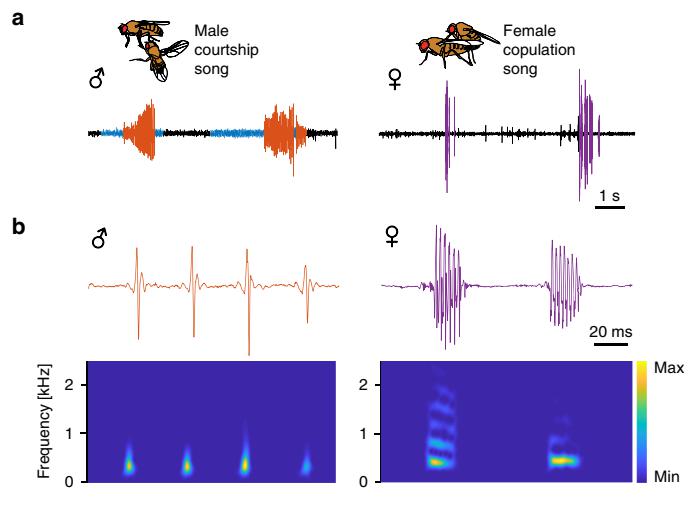
Рис. 2. Песни самца и самки Drosophila.
a Осциллограммы брачной песни самца (импульсная песня показана красным цветом, синусоидальная – голубым) и копуляционной песни самки (пурпурного цвета) с типичными позами во время брачного ритуала.
b Спектрограммы импульсов песни самца и самки.
Возникает резонный вопрос, контролируется ли исполнение песни нервно-мышечным аппаратом самки как это происходит у самцов? Для осуществления крыловой вибрации во время полета и генерации брачной песни требуется активность двигательных нейронов, иннервирующих дорсальные продольные мышцы непрямого действия. Глушение этих нейронов у самок с помощью столбнячного токсина полностью элиминирует песню, тогда как у самца приводит только к уменьшению амплитуды акустического сигнала. Потерю копуляционной песни у самок можно также добиться, заглушив нейроны, экспрессирующие специфический фактор определения пола DoublesexF (dsxF), который к тому же ответственен за репродуктивное поведение самки. Исходя из данных эксперимента по подавлению определенного набора нейронов, авторы предположили, что самкам для генерации песни достаточно активации нейронов брюшной нервной цепи.
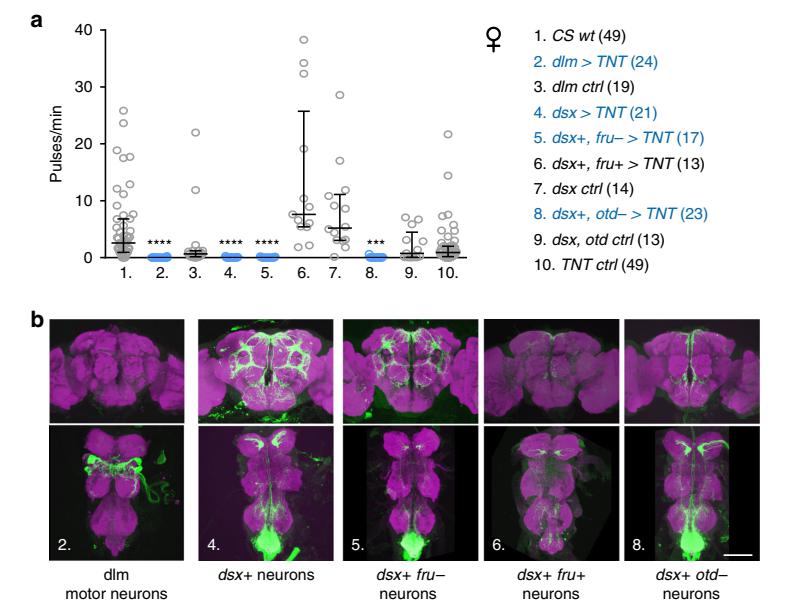
Рис. 3. Нейрональный контроль копуляционной песни самки.
a Количество песенных импульсов на минуту копуляции в контроле и при разных вариантах глушения нейронов с использованием столбнячного токсина.
b Нервная система дрозофилы, состоящая из мозга и брюшной нервной цепи и паттерн экспрессии в структурах нервной системы.
Но что является триггером, запускающим акустическое поведение у самки во время спаривания? Какие сенсорные стимулы вызывают ее пение и оказывает ли влияние спаривающийся самец? Известно, что во время копуляции вместе с эякулятом осуществляется перенос половых пептидов, выделяемых мужскими придаточными железами, что приводит к снижению либидо самки и, как следствие, к снижению вероятности повторных спариваний с другими самцами. Еще одним примером адаптации, повышающим репродуктивный успех самца, является так называемое стратегическое распределение эякулята, что предполагает ценность и быструю истощаемость данного ресурса. Иными словами, самец может контролировать количество, качество и состав семенной жидкости в зависимости от присутствия конкурентов и брачного статуса самки (виргинная или спарившаяся). Это является классическим примером конфликта полов; и какие защитные контрадаптации удалось выработать самкам в этой ситуации, пока доподлинно не известно. Но тем не менее, установленным фактом является то, что самка может довольно быстро спариться со вторым самцом и то, насколько быстро это произойдет зависит от переданного ей состава эякулята первого самца.
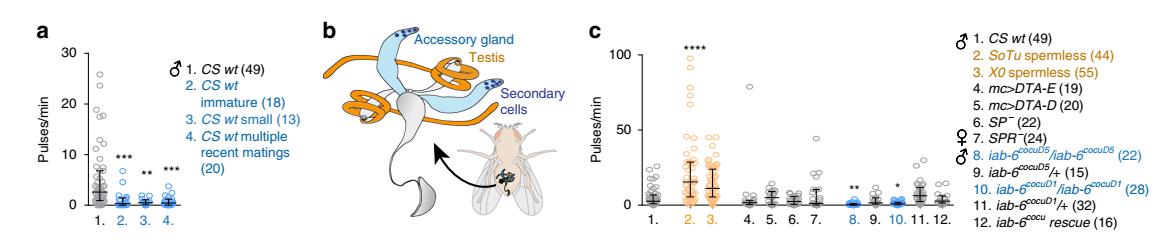
Рис. 4. Зависимость копуляционной песни самки от передачи семенной жидкости самца.
a Количество импульсов копуляционной песни самок дикого типа на минуту копуляции в парах с самцами дикого типа при различных условиях.
b Схема внутренних репродуктивных органов самца. Оранжевым цветом проиллюстрированы семенники, синим – придаточные железы.
c Количество песенных импульсов на минуту копуляции (самки дикого типа и самки-мутанты по SPR рецептору в парах с самцами с нарушенной выработкой сперматозоидов или недостаточной секрецией придаточных желез). Увеличение частоты импульсов песни показаны оранжевым цветом, снижение – синим.
Длительность копуляции у D. melanogaster составляет примерно 15-20 минут, в частности, перенос семенной жидкости осуществляется в течение всего времени, тогда как на перенос сперматозоидов отводится всего 1 минута спустя 7-8 минут от начала спаривания. Акустическое поведение самок во время копуляции зависит не столько от предшествующего спариванию ухаживания, сколько от состава эякулята самца. Чаще всего самки поют в начале и в конце копуляции. Отсутствие спермы в семенной жидкости приводит к увеличению частоты импульсов песни самок. Самцы, испытывающие недостаток эякулята или продуктов вторичных клеток, практически не удостаивались пения самки.
Авторы выдвинули гипотезу, что пение самки в начале спаривания сигнализирует о ее реакции на компоненты семенной жидкости, а пение в конце копуляции стимулирует самца передать ценные компоненты эякулята, необходимых для удовлетворения физиологических потребностей самки. В случае достижения желаемого, повторное спаривание происходит гораздо позже, после эффективного использования материала, полученного от первого самца.
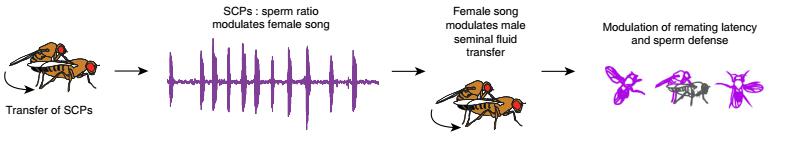
Рис. 5. Предполагаемая схема диалога брачных партнеров во время копуляции.
Еще одним камнем преткновения, усугубляющим конфликт интересов полов, является длительность копуляции. Известно, что у D. melanogaster продолжительность копуляции всецело определяется самцом, тогда как у видов группы virilis она регулируется обоими полами через их взаимодействие между собой. Можно предположить, что появление копуляционной песни у самок необходимо для обеспечения обратной связи между партнерами. Резюмируя написанное выше, можно сказать, что генерация акустического сигнала самкой во время копуляции может быть той самой контрадаптацией, которая позволяет самке искусно манипулировать самцом для извлечения максимальной выгоды.
Новость подготовилa © Белкина Е.Г.
01.06.2020